
Abstract
Microbial composition and functions in the rhizosphere—an important microbial hotspot—are among the most fascinating yet elusive topics in microbial ecology. The aim of this study was to isolate and characterize culturable rhizospheric bacterial isolates from the rhizosphere soil of Aloe barbadensis (dry soil) and Ocimum basilicum (wet soil), focusing on their morphological and biochemical traits. A total of 6 bacterial isolates were purified and screened for PGP characteristics. Morphological characterization included colony color, configuration, size and elevation, while cellular morphology was assessed using simple, Gram, and endospore staining techniques. Biochemical tests (indole, catalase, oxidase) and IAA production in tryptophan-supplemented medium were performed. All isolates were cream-colored but differed in configuration and elevation. Notably, IAA production ranged from 2.1 to 5.4 µg/mL. The isolates displayed diverse biochemical traits, suggesting adaptability to different rhizosphere environments. These findings support the potential of rhizospheric bacteria in sustainable agriculture, particularly in plant growth promotion and stress mitigation.
Introduction
The rhizosphere is the narrow region of soil directly influenced by plant root exudates and associated microorganisms1. It is a dynamic environment where complex interactions occur between plant roots, soil, and microbes, significantly impacting plant health and soil fertility. Root exudates, including sugars, amino acids, and organic acids, stimulate microbial growth and activity, making the rhizosphere a microbial hotspot for nutrient cycling and signaling2.
Rhizospheric bacteria play crucial roles in maintaining soil health and promoting plant growth through various mechanisms. These include decomposition of organic matter, nitrogen fixation, phosphorus solubilization, and the production of phytohormones such as auxins, cytokinins, and gibberellins3,4. In addition, these microbes contribute to plant defence by producing antibiotics or by outcompeting harmful pathogens5.
With rising concerns over the environmental impact of synthetic agrochemicals, plant growth-promoting rhizobacteria (PGPR) offer an eco-friendly alternative, functioning as biofertilizers and biocontrol agents6. Studies have documented the beneficial effects of PGPR in crops such as wheat, maize, and peas grown under various environmental conditions7,8.
Numerous studies have highlighted the beneficial effects of rhizospheric bacteria on various crops. For example, research on the native plant growth-promoting (PGP) bacteria from wheat rhizosphere in the Himalayan region of Rawalkot, Azad Jammu and Kashmir, Pakistan, demonstrated significant benefits for early wheat growth9. Similarly, studies on organic pea grown in Sikkim, India, identified bacterial isolates with high PGP activity, showcasing their potential as biofertilizers10. The characterization of rhizosphere bacteria from maize plants irrigated with industrial and municipal wastewater also revealed significant resistance to heavy metals and salinity, underscoring their utility in bioremediation11.
In this study, Aloe vera and Ocimum basilicum (Basil) were chosen as source plants due to their contrasting rhizospheric environments—Aloe vera is adapted to arid conditions, while Basil prefers moist environments. This selection enabled a comparison of bacterial communities adapted to different moisture regimes. Both plants are also relevant due to their use in herbal medicine and widespread cultivation. The objective was to isolate and characterize rhizospheric bacteria from these two plants, assess their morphological and biochemical traits, and evaluate their potential as PGPR via indole-3-acetic acid (IAA) production. By analyzing culturable strains from ecologically distinct niches, this study contributes to our understanding of plant-microbe interactions and their applications in sustainable agriculture.
Materials and Methods
Sampling from Two Sites
Soil samples were collected from two sites: the dry soil of a flower pot containing Aloe vera (S1) and the wet soil of a Basil plant (S2). One gram of each sample was weighed and stored using a sterile spatula.



Culture Media Preparation
Nutrient agar was prepared using 0.5 g peptone, 0.3 g beef extract, 0.5 g NaCl, and 1.7 g agar in 100 mL of distilled water. The pH of the medium was adjusted to 6.8 by carefully adding 1N NaOH or 1N HCl, and verified using a calibrated digital pH meter. Nutrient agar was selected for its broad utility in cultivating a diverse range of non-fastidious heterotrophic bacteria. Its non-selective nature made it ideal for preliminary isolation before application of differential media in subsequent studies.
Autoclaving
All media were autoclaved at 121°C for 15 minutes prior to use. Sterility was maintained throughout the procedure using a laminar airflow hood, sterile Petri dishes, and autoclaved glassware.

Serial Dilution
A 10-fold serial dilution method was used to reduce microbial concentration prior to plating. One gram of rhizospheric soil was added to 9 mL of sterile distilled water in a test tube and vortexed thoroughly to create a homogeneous soil suspension (10⁻¹ dilution). From this suspension, 1 mL was transferred to a fresh tube containing 9 mL of sterile distilled water to make the 10⁻² dilution, and the process was repeated up to 10⁻⁶. Dilutions were conducted between flames to maintain sterility. Aliquots from selected dilutions (10⁻⁴ to 10⁻⁶) were plated for bacterial isolation.

Inoculation and Incubation
Approximately 15–20 mL of molten nutrient agar, cooled to ~45°C, was poured into each plate. The plates were gently swirled to ensure even distribution of the sample throughout the medium. This pour plate method allowed for the growth of both surface and embedded colonies. Plates were allowed to solidify and incubated in an inverted position at 37°C for 24–48 hours. All inoculations were performed inside a laminar flow hood using sterile pipettes and spreaders to minimise contamination.
Plate preparations were performed in triplicate for each dilution to ensure reproducibility and allow for mean colony count estimation.
Streaking
To isolate pure colonies, streaking was performed using the following procedure: A sterile nichrome loop was used for streaking. The loop was sterilized by passing it through a flame until red-hot and then allowing it to cool. A small amount of the bacterial sample was picked up with the loop and streaked back and forth across the first quadrant of the agar plate to create a dense streak of bacteria. The loop was sterilized again. After cooling, it was used to make several streaks from the edge of the first streak into the second quadrant, spreading the bacteria further. The loop was sterilized once more and used to streak from the edge of the second streak into the third quadrant, further diluting the bacteria. Finally, the loop was sterilized, and a few streaks were made from the third quadrant into the fourth quadrant, ideally resulting in isolated colonies. The plates were incubated inverted at 37°C for 24-48 hours to allow colony formation.
Staining Procedures
Simple staining was performed by preparing bacterial smears, heat-fixing them, and staining with crystal violet. For Gram staining, the standard procedure was followed, including the application of crystal violet, Gram’s iodine, ethanol decolorizer, and safranin counterstain. Schaeffer-Fulton endospore staining involved applying malachite green, steaming, and counterstaining with safranin. The hanging drop method was used to observe bacterial motility by placing a drop of bacterial suspension on a coverslip, sealing it with Vaseline, and inverting it onto a concavity slide.

Biochemical Tests
Indole test: The indole test was conducted to determine the ability of bacterial isolates to produce indole from tryptophan, indicating the presence of the enzyme tryptophanase. Each isolate was inoculated into tryptone broth and incubated at 37°C for 24–48 hours. After incubation, 0.5 mL of Kovac’s reagent (para-dimethylaminobenzaldehyde in amyl alcohol and hydrochloric acid) was added. Tubes were gently shaken and observed for the formation of a red or pink layer at the top, indicating a positive result.
Catalase test: This test detects the presence of the catalase enzyme, which breaks down hydrogen peroxide (H₂O₂) into water and oxygen. A small amount of bacterial colony was transferred onto a clean, grease-free glass slide, and a drop of 3% hydrogen peroxide was added. The immediate appearance of bubbles indicated a positive result.
Oxidase test: The oxidase test was used to detect the presence of cytochrome c oxidase. A bacterial sample was transferred onto an oxidase test disc using a sterile loop. A color change from white to blue or purple within 10–30 seconds was considered a positive result.
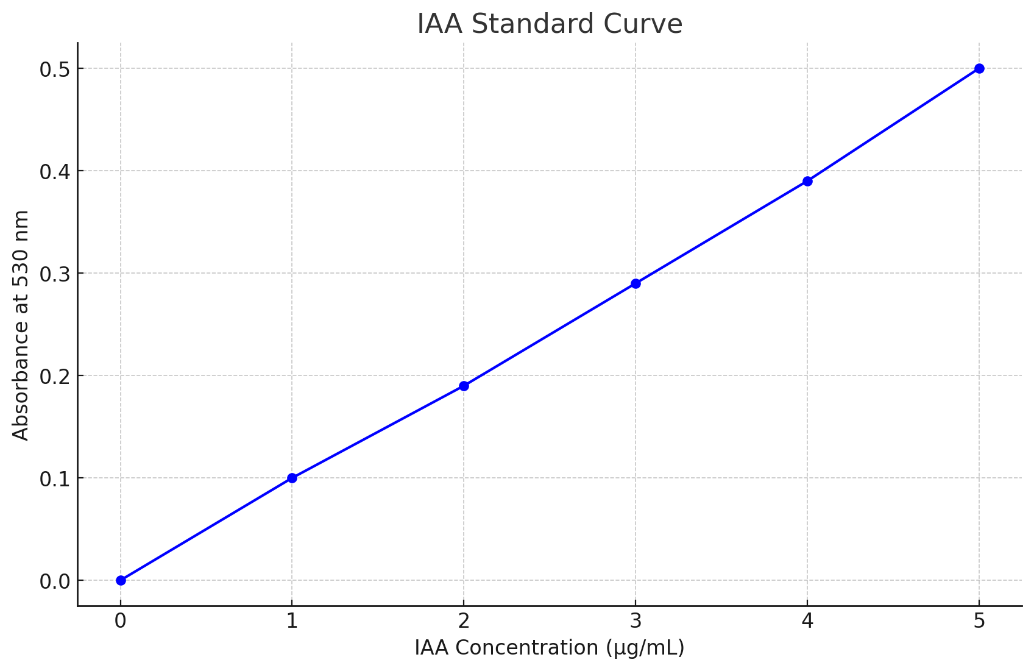
IAA quantification: The ability of bacterial isolates to produce indole-3-acetic acid (IAA) was assessed using the Salkowski colorimetric method. Each isolate was grown in nutrient broth supplemented with 0.1% L-tryptophan and incubated at 30°C for 48 hours under shaking conditions. After incubation, cultures were centrifuged at 10,000 rpm for 10 minutes. One milliliter of the supernatant was mixed with 2 mL of freshly prepared Salkowski reagent (1 mL of 0.5 M FeCl₃ in 50 mL of 35% perchloric acid). The mixture was incubated in the dark at room temperature for 30 minutes. Development of a pink color indicated IAA production. Absorbance was measured at 530 nm using a spectrophotometer, and IAA concentration (µg/mL) was calculated using a standard curve generated from known concentrations of pure IAA.
All biochemical tests and IAA quantification assays were conducted in triplicate. Results were recorded as consistent if at least two of the three replicates yielded the same outcome. For IAA production, the average absorbance value from three trials was used to estimate final concentrations via the standard curve.
All incubations for biochemical tests were conducted at 37°C, with durations ranging from 24 to 48 hours depending on the test. Sterile conditions were maintained by using pre-autoclaved media and performing inoculations under aseptic conditions.
Results and Discussion
Isolation and Characterization of Bacteria
Bacterial colonies were successfully isolated from both soil samples (S1 and S2).

While most further isolates initially appeared cream-colored on nutrient agar, closer observation revealed subtle variations in hue ranging from off-white to light beige. Differences in surface texture and margin sharpness were also noted. Colonies displayed irregular or rhizoid configurations, with irregular colonies having uneven, wavy edges and rhizoid colonies having root-like branching structures. The colonies were observed to have either flat or raised elevations.

The average colony diameter across all isolates ranged from 1.8 mm to 3.2 mm after 48 hours of incubation at 37°C. Colonies from Aloe vera samples tended to be slightly larger, with S1-A1 reaching 3.2 mm, while Basil-derived isolates generally ranged between 1.8 mm and 2.5 mm. All measurements were taken in triplicate using a digital Vernier caliper, and the mean value was reported.
Staining Results
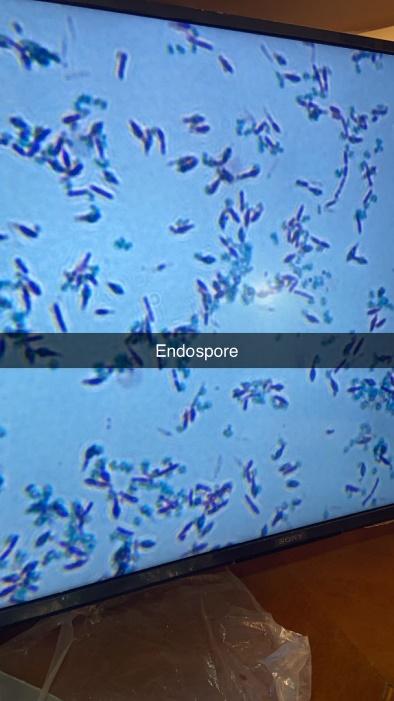
Simple staining revealed various shapes and arrangements of bacterial cells. Gram staining identified both Gram-positive (dark purple) and Gram-negative (pink) bacteria among the isolates: out of the six, three were Gram-positive and three were Gram-negative. Schaeffer-Fulton endospore staining showed endospores as green, with the rest of the cell appearing pink under oil immersion (100x) objective. The hanging drop method indicated non-motile bacteria, as no movement was observed.

Biochemical Tests
Biochemical characterization of the isolates included tests for catalase, oxidase, indole production, and IAA quantification. All six isolates tested positive for catalase activity, indicating their ability to degrade hydrogen peroxide into water and oxygen, a trait linked to oxidative stress tolerance. Oxidase activity was negative in all isolates, suggesting the absence of cytochrome c oxidase, and indicating that these strains are unlikely to belong to genera such as Pseudomonas.
Indole production varied across isolates, with four out of six testing positive, as indicated by the development of a red-pink layer upon addition of Kovac’s reagent. The remaining two isolates showed no color change.
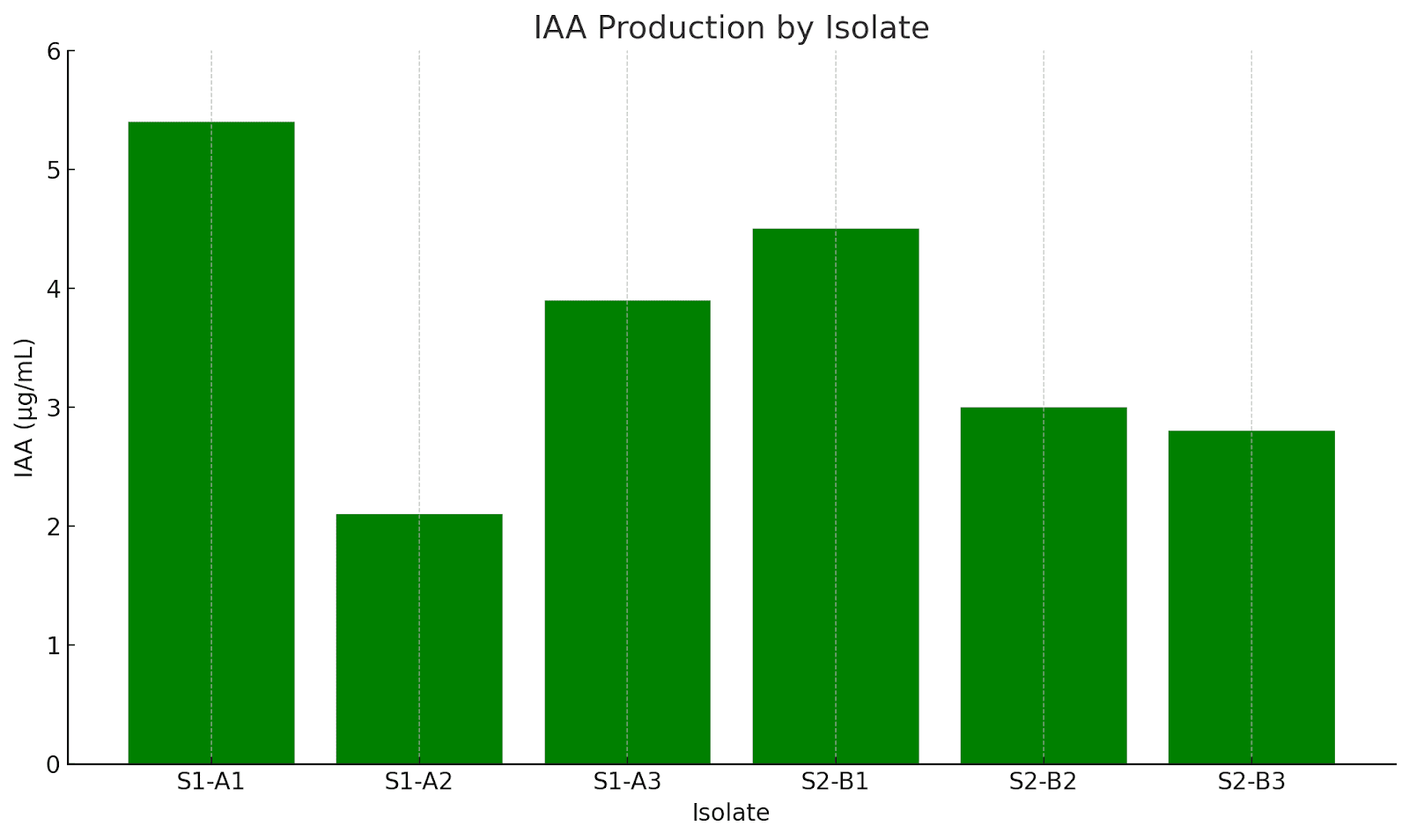
IAA quantification was performed using the Salkowski reagent assay. Isolates were grown in tryptophan-supplemented broth, and absorbance was measured at 530 nm. IAA production values represent the mean of three replicates. The highest-producing isolate (S1-A1) yielded 5.4 µg/mL (±0.2), while the lowest (S1-A2) produced 2.1 µg/mL (±0.1).. All values were determined based on a standard curve derived from known concentrations of pure IAA, depicted below.
The results of all biochemical tests, along with morphological and staining data, are summarized in Table 1.
| Isolate | Source Plant | Colony Morphology | Gram Stain | Endospore | Indole | Catalase | Oxidase | IAA (µg/mL) |
| S1-A1 | Aloe vera | Cream, Irregular, Raised | + | + | + | + | – | 5.4 |
| S1-A2 | Aloe vera | Cream, Rhizoid, Flat | – | – | – | + | – | 2.1 |
| S1-A3 | Aloe vera | Cream, Circular, Raised | + | + | + | + | – | 3.9 |
| S2-B1 | Ocimum basilicum | Cream, Irregular, Flat | – | – | + | + | – | 4.5 |
| S2-B2 | Ocimum basilicum | Cream, Rhizoid, Raised | + | – | – | + | – | 3.0 |
| S2-B3 | Ocimum basilicum | Cream, Circular, Flat | – | – | – | + | – | 2.8 |
Discussion
The isolates demonstrated a variety of biochemical and morphological traits, indicating their adaptability to distinct rhizospheric conditions. Gram staining revealed that out of the six isolates, three were Gram-positive and three were Gram-negative. Notably, Gram-positive isolates were more frequent in the dry soil of Aloe vera, suggesting a potential link between cell wall structure and desiccation resistance12. Two endospore-forming strains were also found exclusively in Aloe vera samples, which may offer these bacteria an ecological advantage under drought conditions, as endospores allow long-term survival during nutrient limitation or desiccation4. These structures offer a survival advantage under desiccation and nutrient-limited conditions, suggesting a well-adapted rhizobacterial population evolved to persist in xeric environments. This aligns with the broader ecological role of Bacillus spp. in arid soils13. The relatively larger colony sizes observed in Aloe vera isolates may also indicate a more aggressive growth phenotype, possibly linked to the aforementioned adaptations.
All isolates (100%) tested catalase-positive, indicating their ability to detoxify reactive oxygen species—a crucial trait in rhizosphere survival14. Such antioxidant defences are particularly valuable under stress conditions like drought or high root exudate activity, where oxidative bursts can threaten microbial persistence15. The oxidase test was negative in all isolates (0%), suggesting the absence of cytochrome c oxidase and ruling out oxidase-positive genera such as Pseudomonas. Instead, the traits observed align with genera like Bacillus, Azospirillum, or Enterobacter—well-known for their plant-associated functions2.
Indole production results were variable. Four isolates produced indole (66.7%), as confirmed by colour change with Kovac’s reagent, while two did not. Quantitative IAA analysis revealed concentrations ranging from 2.1 to 5.4 µg/mL, with isolate S1-A1 from the Aloe vera rhizosphere producing the highest amount. Previous studies suggest that IAA concentrations above 5 µg/mL may significantly enhance root elongation and cell division, indicating the potential of such isolates as strong PGPR candidates16. This trait could be especially beneficial under nutrient-limited conditions, as IAA-producing microbes often improve nutrient uptake through root surface expansion.
The contrast in colony configurations, Gram nature, and endospore presence between isolates from different plant hosts supports the notion that abiotic factors like soil moisture shape bacterial trait evolution and community composition. The dominance of endospore-formers in the dry Aloe vera rhizosphere supports stress adaptation, while the prevalence of Gram-negative, non-spore-forming strains in Basil suggests an environment favouring faster growth but less structural resistance. These observations reflect classic microbial ecological strategies in response to soil physicochemical conditions.
The comprehensive profiling of each isolate—including staining behaviour, morphology, and metabolic traits—was summarised in Table 1. Together, these findings underscore the ecological and biotechnological significance of rhizobacteria and suggest that further investigation using molecular identification (e.g., 16S rRNA or RFLP) and greenhouse trials is warranted to confirm their plant growth-promoting capabilities and stress resilience. Future work should also include antimicrobial assays and growth kinetics under varying environmental conditions. Such analyses would further elucidate the functional roles of these isolates.
Conclusion
This study successfully isolated and characterized culturable bacterial strains from the rhizosphere soil of Aloe vera and Basil plants. The isolates exhibited diverse morphological traits and significant biochemical capabilities, such as indole production, catalase activity, and oxidase negativity. These traits highlight the potential of these rhizospheric bacteria in promoting plant growth and maintaining soil health, making them valuable for sustainable agricultural practices.
Further studies using molecular techniques such as 16S rRNA gene sequencing, PCR-based fingerprinting, or RFLP, depending on resource availability, are necessary to identify the bacterial strains and understand their phylogenetic relationships. Assessing the specific biochemical pathways and genes involved in indole production and other plant growth-promoting activities can provide deeper insights into their functional roles. Conducting field trials to evaluate the efficacy of these bacterial isolates in enhancing crop yield and health under natural conditions will be crucial for practical agricultural applications.
References
- B. Lugtenberg, F. Kamilova, Plant-growth-promoting rhizobacteria. Annual Review of Microbiology 63, 541–556 (2009). [↩]
- R. Mendes, P. Garbeva, J. M. Raaijmakers, The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiology Reviews 37, 634–663 (2013). [↩] [↩]
- B. R. Glick, Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 1–15 (2012). [↩]
- R. Hayat, S. Ali, U. Amara, R. Khalid, I. Ahmed, Soil beneficial bacteria and their role in plant growth promotion: A review. Annals of Microbiology 60, 579–598 (2010). [↩] [↩]
- S. Compant, C. Clément, A. Sessitsch, Plant growth-promoting bacteria in the rhizo- and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biology and Biochemistry 42, 669–678 (2010). [↩]
- P. N. Bhattacharyya, D. K. Jha, Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World Journal of Microbiology and Biotechnology 28, 1327–1350 (2012). [↩]
- J. K. Vessey, Plant growth promoting rhizobacteria as biofertilizers. Plant and Soil 255, 571–586 (2003). [↩]
- F. Liaqat, R. Eltem, Isolation and characterization of potent phosphate solubilizing bacteria from plant rhizosphere. Frontiers in Microbiology 12, 733027 (2021). [↩]
- A. Tkacz, J. Cheema, G. Chandra, A. Grant, P. S. Poole. Stability and succession of the rhizosphere microbiota depends upon plant type and soil composition. The ISME Journal, 9, 2349–2359 (2015). [↩]
- E. Deyett, C. Roper, P. Rolshausen. Microbial community profiling of grapevine xylem microbiome using nanopore sequencing. Current Research in Microbial Sciences, 2, 100049 (2021). [↩]
- M. Abedinzadeh, H. Etesami, H. A. Alikhani. Characterization of rhizosphere and endophytic bacteria from roots of maize (Zea mays L.) plant irrigated with wastewater with biotechnological potential in agriculture. Biotechnology Reports, 21, e00305 (2019). [↩]
- R. Backer, J. S. Rokem, G. Ilangumaran, J. Lamont, D. Praslickova, E. Ricci, N. Subramanian, D. L. Smith, Plant growth-promoting rhizobacteria: Mechanisms and applications. Scientifica 2018, 1–15 (2018). [↩]
- S. S. Singh, M. Singh, B. S. Saharan, Microbial endophytes: Potential biocontrol agents for managing plant diseases. Biological Control 144, 104236 (2020). [↩]
- D. Egamberdieva, S. J. Wirth, A. A. Alqarawi, E. F. Abd_Allah, A. Hashem, Phytohormones and beneficial microbes: Essential components for plants to balance stress and fitness. Frontiers in Microbiology 8, 2104 (2017). [↩]
- D. K. Maheshwari, R. Dheeman, B. Agarwal, Phytohormone-producing PGPR for sustainable agriculture. In: D. K. Maheshwari (ed.) Bacterial Diversity in Sustainable Agriculture, Springer, 159–182 (2014). [↩]
- S. Patten, B. R. Glick, Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Applied and Environmental Microbiology 68, 3795–3801 (2002). [↩]


