
Abstract
Gestational diabetes mellitus (GDM) affects the maternal gut microbiome, influencing fetal development and contributing to congenital fetal abnormalities, such as cleft palate defects. GDM is a condition in which a woman develops high blood sugar levels during pregnancy due to dysfunctional pancreatic β-cells. This condition alters the pregnant women’s ever-evolving gut microbiome, which is critical in maintaining overall health. Changes in the microbiome due to GDM can disrupt key developmental processes in the fetus, creating an environment that can contribute to orofacial abnormalities. Notably, GDM promotes microbiome imbalances that increase inflammatory bacteria, such as Desulfovibrio, while also elevating Lactobacillus levels in the placental microbiome—bacteria associated with cleft-affected newborns. Lactobacillus is associated with cleft-affected newborns and has been tied to dysregulations of the Wnt signaling pathway, which plays a critical role in orofacial formation. Furthermore, this paper explores the connection between GDM-induced microbiome changes and orofacial defects, focusing on inflammatory pathways and key developmental signaling mechanisms. GDM enhances bacterial movement into the bloodstream, triggering systemic inflammation that promotes CDH1 hypermethylation, a process linked to impaired neural crest cell migration and craniofacial malformations. Additionally, GDM-induced microbiome alterations lead to disruptions in Wnt signaling, a pathway essential for facial development, further contributing to cleft formation. Beyond microbial influences, GDM is associated with elevated retinoic acid (RA) levels, which can interfere with proper craniofacial development by disrupting key signaling pathways. RA overexpression impairs normal palatal fusion, increasing the risk of orofacial clefts. Moreover, the dysregulation of calcium homeostasis in GDM pregnancies affects the Wnt5a-Ca²⁺ pathway, which plays a critical role in jaw and palate formation. Although cleft palates can be surgically corrected soon after birth, they profoundly impact maturing children, leading to hearing, speaking, breathing, and socialization difficulties. Targeting one potential origin of this defect can increase awareness and potentially prevent future cases.
Introduction
A healthy person contains an average of 39 trillion microbes, microscopic organisms like bacteria, archaea, single-cell eukaryotes, and protozoa. Each person has a unique and original microbiome influenced by genetic and environmental factors1.
For many years, it was believed that a fetus was sterile in utero. However, recent studies show that the composition of one’s microbiome starts before birth. Bacteria found in the placenta, amniotic fluid, and the umbilical cord are the building blocks of the fetus’ microbiome, shaping the immune, digestive, and central nervous systems. As life progresses, additional factors, including lifecycle stages, diet, geography, medications, stress, and substance exposure, can alter the health and regulation of the microbiome. The makeup of the gut microbiome plays a significant role in determining a person’s overall health and well-being2.
The microbiome is also significant during the perinatal period as the fetus’ brain undergoes rapid growth in correspondence to changes in the maternal microbiome. Microbes such as Proteobacteria and Actinobacteria increase from the first to the third trimester of pregnancy. The maternal microbiome further influences fetal brain development through its role in tryptophan metabolism and serotonin synthesis (5-HT). Tryptophan metabolism refers to the process of breaking down the amino acid tryptophan into different substances, and serotonin synthesis is the process of turning tryptophan into serotonin. The maternal microbiome metabolizes tryptophan and synthesizes serotonin, which is taken up by the fetus and used by its developing central nervous system. A study supporting this connection showed that mice with a modified TPH2 gene—where the gene responsible for serotonin (5-HT) production in the brain was replaced by an EGFP reporter gene—exhibited improper brain wiring and potential long-term neurodevelopmental disorders. It was noted that these abnormalities could have been prevented by exposing the mice to gut microbiota early in life, emphasizing how alterations to the maternal microbiome during pregnancy can shape the fetal brain’s development. The study showed that disruptions to the microbiome’s role in metabolic pathways can lead to dysfunctions, highlighting the importance of maintaining a healthy microbiome during pregnancy for optimal fetal health3.
Microbiomes are also vital in protecting and defending the body from infections. Good microbes work to maintain the body’s health, and altercations to them increase the body’s risk of contracting infections4.
Diseases, along with other environmental factors, possess the ability to alter the microbiome. One such disease is GDM, also known as maternal diabetes. Annually, this condition affects more than 200,000 pregnancies in the US and approximately 10-16.5% worldwide, making it one of the most common medical complications in pregnancy. Research associates GDM with changes in the maternal gut microbiome as it decreases microbial diversity and increases the levels of inflammatory microbe bacteria5.
A sudden spike in blood sugar classifies GDM due to the dysfunction of pancreatic β-cells responsible for producing insulin. When these cells malfunction, they fail to produce sufficient insulin to counteract insulin resistance, where the body’s cells become less responsive to insulin, resulting in elevated blood sugar levels. Normally, insulin sensitivity will spike early in gestation to promote the conversion of glucose into fats, which are later used for energy. However, in GDM pregnancies, the increase in insulin sensitivity is impaired, leading to higher blood sugar levels as the body cannot regulate glucose metabolism6.
The systematic differences GDM creates impact not only the pregnant women but also the developing fetus. GDM increases the pregnant women’s and fetus’ risk of developing type II diabetes after birth6 and can contribute to developmental fetal defects such as cleft plates in neonates.
Being the 4th most common congenital malformation, orofacial clefts (OFCs) affect one out of every 650 births. Children born with this condition can present in three ways: cleft palate, cleft lip, or concurrent cleft lip and palate7.
A cleft palate occurs when improper fusion creates a hole or opening in the roof of the mouth, which develops from the fourth to twelfth week of gestation. Before this period, embryonic folding leads to the formation of the stomodeum, a depression that develops into the mouth. The stomodeum opens into the primitive pharynx, an embryonic structure that will help create the throat. Within the primitive pharynx, the pharyngeal arches are formed from six thickenings. These thickenings will help develop into embryonic connective tissues through the help of neural crest cells (NCC) and are used later for facial formation. This process is complete when the maxilla and palatine bones fuse with the palatine process, which originates from one of the six arches, to form the hard palate. Dysfunction of these processes results in cleft palates. A cleft lip is a split or opening in the upper lip resulting from a lip closure dysfunction. Development occurs as the maxillary prominences fuse with the lateral nasal prominence to form parts of the upper lip. This process continues with further growth and fusion of the medial nasal prominence, contributing to the formation of the nostrils. Fusions between these prominences result in the formation of mesenchymal tissue, which separates the upper lip from the nostrils and forms the primary palate. A one-sided dysfunction between the maxillary and nasal prominence will result in a one-sided cleft lip. A bilateral dysfunction between the maxillary and nasal prominences will result in a bilateral cleft lip8.
Concurrent cleft lip and palate are commonly presented, but all types of cleft defects (Figure 1, lower panel) create difficulties for the patient. A cleft disrupts the fusion of muscles, affecting speech and swallowing9. Patients typically also suffer from ear infections, dental problems, and social challenges. Surgical procedures are currently the only treatment option10.
This paper will discuss the effect of GDM on the maternal microbiome and how it can lead to cleft palate defects. This question is becoming increasingly relevant as the number of GDM pregnancies rises. Understanding the potential risks and complications associated with GDM is crucial for educating expecting families and the general public about the conditions’ impact on the microbiome and fetal orofacial development11.
Methods
Research for this paper was conducted mainly through platforms such as Google Scholar and PubMed. Keywords such as “gut microbiome,” “gestational diabetes mellitus and microbiome,” “cleft and microbiome,” and “gestational diabetes mellitus and cleft” were used to identify relevant peer-reviewed primary scientific literature and review articles. In some cases, different articles used different terms to explain the same idea; synonyms were also used as keywords. For example, both “gestational diabetes” and “maternal diabetes” were keywords, as well as “orofacial cleft” and “cleft lip/palate.”
Section I: Maternal and fetal microbiome changes associated with GDM
GDM has immense power over the overall health and condition of the human body. GDM affects glucose uptake and metabolism and also works to alter microbiome composition and function.
The most commonly observed difference between the microbiome of an unaffected pregnancy and the microbiome of GDM-affected and non-affected pregnant women is the increase in the Firmicutes group of microbes, along with a decrease in the Bacteroidetes and Actinobacteria groups in the GDM-affected group. Firmicutes, made up of Ruminococcus, Clostridium, Lactobacillus, and butyrate-producing bacteria, is one of the five major bacterial species found in the microbiome. Bacteroidetes are another significant bacteria grouping12. Firmicutes and Bacteroidetes comprise more than 90% of the gut microbiome; the Firmicutes/Bacteroidetes ratio indicates overall health and positively correlates with reduced glucose tolerance13. In addition, neonates born to women with GDM have been found to have high concentrations of Lachnospiraceae, a microbe that is also elevated in women with GDM. Shifts in microbiome composition can have long-term effects on maternal and neonatal health, as the presence of Lachnospiraceae could contribute to metabolic disturbances, influencing the development of insulin resistance in both the postpartum women and her neonate1. Another study found several more differences in the microbiome of a GDM-affected woman. Decreased levels of Pseudomonadales, Dialister, Akkermansia, Roseburia, Bacteroides, Methanobrevibacter smithii, Eubacterium, Alistipes, Bifidobacterium and Lactobacilli, increased levels of Firmicutes, Klebsiella variicola, Collinsella, Rothia, Ruminococcus, Actinobacteria, Parabacteroides distasonis, and Desulfovibrio were seen in GDM-affected women compared to non-affected pregnant women14. Interestingly, changes in the gut microbiome did not always correspond to changes in the oral, vaginal, or placental microbiomes. Specifically, Lactobacillus—a bacteria commonly elevated in children with cleft palate—was increased in the placental microbiome but decreased in the gut and oral cavity microbiome of GDM-affected women compared to non-affected pregnant women.
Changes in the maternal microbiome can affect the fetus during development and have long-term postnatal impacts, even as long as 16 months after birth. Lower levels of Erysipelotrichaceae, Flavonifractor, Lactobacillus, and Gammaproteobacteria in the maternal gut microbiome, for example, are associated with higher concentrations of these microorganisms within the baby. GDM changes in the maternal microbiome change the fetus by indirectly altering the fetal microbiome. Altercations to the fetal microbiome are consequential as microbes are responsible for early immune cell function suppression. This affects how immune cells respond to infections, recognize pathogens, and develop long-term immunity, changes that affect overall fetal health and development1.
A study at the Women’s Hospital of Nanjing Medical University supported the impact of GDM on a baby’s microbiome. After birth, the babies’ first stool samples were collected and tested. The results revealed a noticeable decrease in the alpha(α)- diversity of babies born to GDM-affected women compared to those born to unaffected women. α-diversity refers to species variation within a given area; in this case, babies born from GDM pregnancies exhibited less microbial diversity. A decrease in microbiome diversity is concerning because it can compromise the infant’s immune system development, increase susceptibility to infections and diseases, and affect the ability to digest and absorb nutrients effectively. Diminished diversity is particularly significant as it may also impact fetal development, potentially contributing to congenital defects such as cleft palate.
Additionally, the study found that GDM is linked to shifts in beta(β)- diversity levels, which refers to the similarity of microbes within a given area15. Disruptions in both α- and β-diversity could influence the baby’s immediate health and may have long-term implications, including the development of conditions like cleft defects in utero, as the altered microbiome may affect critical developmental processes. Interestingly, the β-diversity levels also differed due to delivery modes, further highlighting how the maternal microbiome influences the fetus. Since different microbiome systems contain different microbes, the baby acquires microbes based on its exposure during birth. Compared to the control group of babies born to normal pregnancies, babies born to women with GDM had an abundance of Firmicutes16. Women with GDM also displayed higher levels of Firmicute, as well as Collinsella, Rothia, Desulfovibrio, Actinobacteria, Parabacteroides distasonis, Ruminococcus, Eubacterium, and Prevotella17. Elevated levels of bacteria, such as previously listed, can be related to orofacial or fetal development issues through various mechanisms. For instance, bacteria that promote inflammation, such as Desulfovibrio, might cause systemic inflammation and disrupt normal fetal development, leading to potential anomalies. An overabundance of bacteria like Ruminococcus, which aids in nutrient breakdown, could alter the balance of nutrient absorption, leading to deficiencies or excesses of nutrients. Nutrients, such as folate and vitamin A, are crucial for proper craniofacial development. Disruptions in nutrient absorption can contribute to cleft defects by impairing the normal development of the facial structures. Additionally, GDM-induced changes in the microbiome can affect the maternal immune system and hormonal balances, both of which are critical for normal fetal development: such imbalances increase the risk of cleft palate.
Building on the consequences of altered microbiomes, recent studies have found that gut microbiome changes can increase intestinal permeability, meaning that bacteria can more readily enter the bloodstream14. Furthermore, additional studies have shown that bacteria from a gravid female can pass through her placenta and enter the fetus’s bloodstream18. Bacteria affect the embryo by influencing immune responses, nutrient absorption, and development. GDM creates proinflammatory environments because higher blood glucose levels stimulate the production of cytokines and other inflammatory markers. These molecules trigger inflammation; elevated inflammatory markers can impair organ development and affect blood flow to the placenta, reducing nutrient and oxygen delivery to the fetus, which increases susceptibility to complications like cleft defects19.
Section II: The Microbiome’s Influence on the Development of Clefts
A study found hyperglycemic rats to have altered levels of inositol and prostaglandins in the gut. These factors affect the development of neural crest-deriving organs, such as the mandible and Meckel’s cartilage. The elevated glucose levels in the rats affected their microbiomes, leading to these developmental issues. The hyperglycemia in GDM could cause similar disruptions in the microbiome of affected women, potentially leading to developmental problems in the mandible and Meckel’s cartilage. Since cleft palate development is linked to defects in these structures, GDM can be tied to the formation of cleft palates.
A study conducted at Marmara University and Research Hospital NewBorn Clinic took intraoral samples from a cleft palate group of twenty-one children and a control group of twelve children. Samples were taken at least three times, starting at birth and ending after three years. Compared to the control group, the level of Lactobacilli bacteria within GDM-born babies was higher at birth (p=0.029) and at the growth of the first primary tooth (p=0.030)20.
An additional study supported these findings through research on eighty five-year-old children in a cleft palate group and 144 age-matched children in a non-cleft palate group. Stimulated saliva samples were collected and analyzed for similarity between each group. The results found higher numbers of Lactobacillus in the cleft palate group20. These studies concluded an association between cleft palate and high levels of Lactobacillus. Despite some varying results, several studies have found higher levels of Lactobacillus in the maternal microbiome of GDM-affected women.
Retinoic acid (RA) is needed for growth, immune response, and embryonic development. In the early stages of development, RA is needed to properly form the heart, hindbrain, spinal cord, and eyes. It is later required to develop the skeleton, lungs, pancreas, and genitourinary tract21. Elevated glucose levels in GDM affect the maternal microbiome by leading to an overabundance of Lactobacillus. Research has shown that, through gut changes and hyperglycemia changes, GDM encourages the activation of retinoic acid receptors (RARs), which control gene expression linked to immune regulation and cell differentiation.
Additionally, the activity of enzymes involved in retinol metabolism, such as retinol dehydrogenase (RDH), which changes retinol (vitamin A) into retinoic acid, might be affected by insulin resistance and high glucose levels. Because of this, RA can accumulate in the pregnant women and fetus and contribute to the formation of cleft defects22. In addition, high vitamin A levels are thought to be associated with higher RA levels because RA is the primary metabolite of vitamin A. A study at Shunyi District Maternal and Child Health Hospital measured the serum vitamin A concentration of 391 women. Women in the highest quartile of vitamin A levels had 2.85 times higher odds (P=0.042) of developing GDM compared to those in the lowest quartile, and each standard deviation increase in vitamin A levels was associated with a 1.59 times higher risk (P=0.011)23.
RA also functions as a critical regulator of protein factors, like fibroblast growth factor (FGF), sonic hedgehog (SHH), transforming growth factor beta (TGFβ), and Wnt. These factors are essential for orofacial development. As Figure 1 illustrates, GDM leads to epigenetic alterations, disrupting key signaling pathways—including Wnt, TGFβ, SHH, FGF, and RA—necessary for proper orofacial formation. Therefore, conditions like GDM, which can result in excess levels of or the dysfunction of RA, inhibit the proper formation of palate structures, giving rise to cleft defect development.
The National Vital Statistics System of The United States of America found women with GDM to have an increased relative risk of 1.36 (95% CI: 1.30–1.43, P<0.0001) for having babies with orofacial clefts compared to women without GDM. Specifically, the relative risk was 1.28 (P<0.001) for cleft lip with or without cleft palate and 1.40 (P<0.0001) for cleft palate only. A physiological reason for these defects can be Wnt dysfunctions24. Within an embryo, Wnt signaling is needed to direct craniofacial development, and more than twenty Wnt signaling genes are tied to OFCs25. The Wnt signaling pathway begins when a Wnt protein attaches to a receptor on the cell’s surface, which causes a signal transduction pathway. The signal transduction pathway receives the signals at the cell’s surface and triggers specific cellular actions in response. Wnt signaling is essential in regulating stem cell population, embryonic development, cell growth, differentiation, and migration. The expression of Wnt signaling can be related to Lactobacillus, as shown by animal studies. A study on young mice found that microbiome bacteria Lactobacillus rhamnosus GG (LGG) and Butyrate promote bone formation and increase bone density by increasing the expression of Wnt10b in bone marrow26. This study highlights how changes in the gut microbiome, which can be caused by GDM, can alter Wnt signaling pathways and influence bone formation, therefore, explaining how disrupted Wnt signals can contribute to defects such as clefts.
Wnt signaling can be classified into canonical and noncanonical pathways; the noncanonical pathways can be broken down into polarity and the Wnt5a-Ca²⁺ pathways24. The Wnt5a-Ca²⁺ pathway involves calcium signaling in its transduction process. High sugar levels have been found to alter serum calcium homeostasis, with women in the highest third of serum calcium levels having an average fasting glucose concentration of 5.06 ± 0.03 mmol/L, compared to 4.80 ± 0.03 mmol/L in the lowest third27. Impairing the Wnt5a-Ca²⁺ pathway leads to orofacial defects, including cleft palate formation. Furthermore, GDM can lead to the overexpression of Wnt genes Sfrp1 and Dkk1.Essential developmental pathways necessary for craniofacial morphogenesis are inhibited when canonical Wnt signaling is suppressed. During the fusion of the lip and palate, the improper activation of β-catenin prevents it from carrying out its roles in cell migration, proliferation, and differentiation. This disruption interferes with the normal development of these structures, ultimately leading to the formation of a cleft palate24.
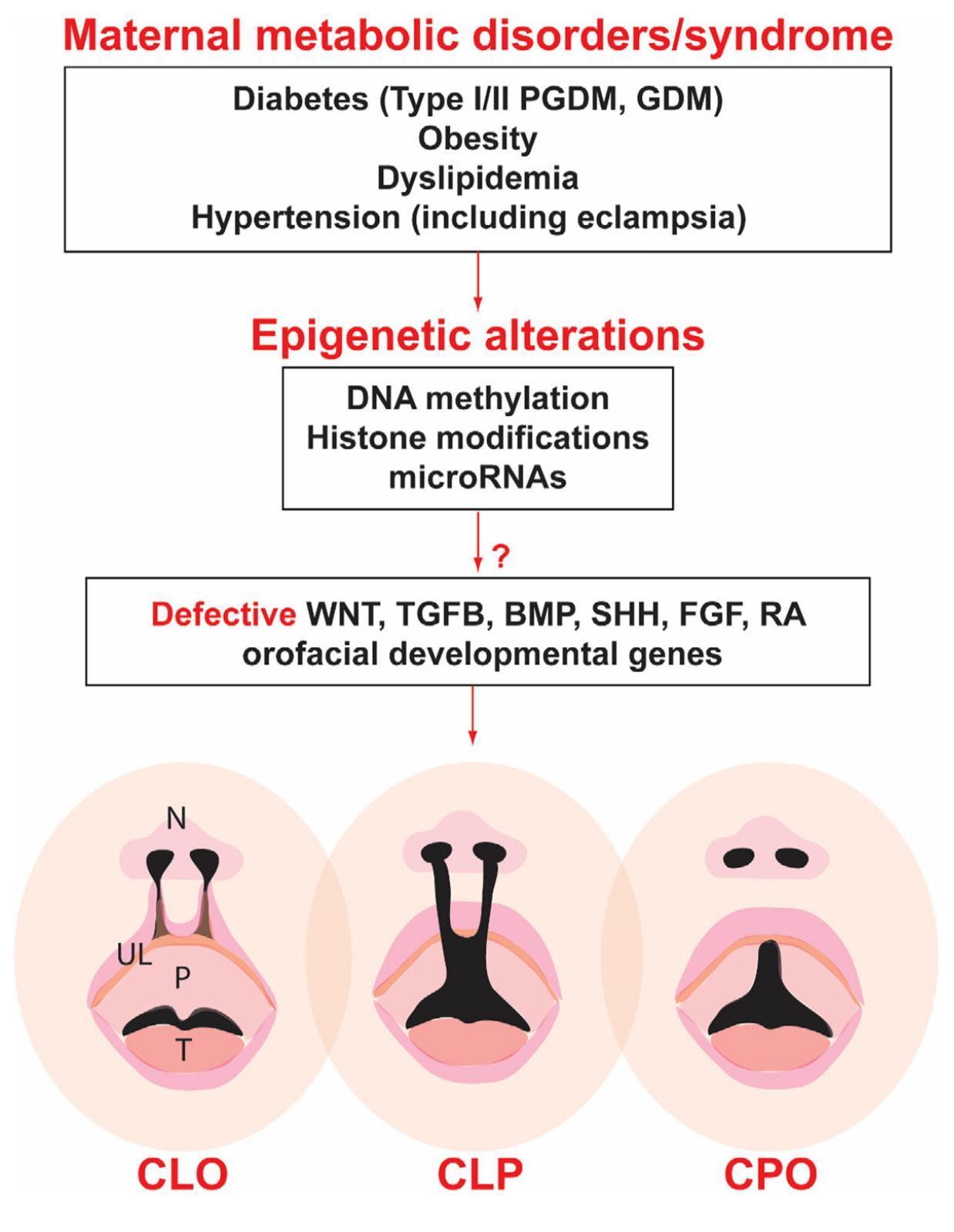
GDM may contribute to the development of cleft palate by allowing bacteria to move from the maternal bloodstream to the fetal circulation, increasing the risk of fetal exposure to microbial factors that could disrupt normal craniofacial development. Studies in mice suggest maternal bacteria can cross the placenta and enter the fetal bloodstream. Fluorescently labeled bacterial cell wall fragments (CW) injected into pregnant mice were detected in the fetal brain after only 24 hours. These findings suggest bacteria from the maternal microbiome can rapidly cross the placenta to enter the fetus. Furthermore, CW is commonly referenced as causing inflammation and cell death, both of which are associated with the development of cleft palates28.
A study analyzing the subgingival oral biofilm, a layer of microbes beneath the gums, of patients with cleft palate found several types of bacteria, including Aggregatibacter, Tannerella forsythia, and Porphyromonas. These patients had a higher gingival bleeding index, plaque index, and clinical attachment level, all of which are indicators of bacterial presence29. Increased bacteria in patients with cleft palates suggests a bacterial link to the abnormality. This relates to GDM because, as previously mentioned, GDM can alter the maternal microbiome and facilitate faster bacterial movement into the bloodstream. This enables maternal bacteria to cross the placenta, and pathogens that can do so can influence fetal development by increasing the bacterial presence. A study infected pregnant mice with bacterial cell wall fragments (CW) and found that the CW had accumulated within the fetal brain only 24 hours later.
Cytokines, inflammatory molecules secreted by immune system cells, cause the hypermethylation of CDH1. CDH1 is the gene that encodes for the protein E-cadherin, which is essential for proper NCC migration. NCCs facilitate embryonic development and, therefore, relate to cleft palate/lip development. Recent studies have found that when both copies of the CDH1 gene are turned off, either by mutation and/or hypermethylation of the gene promoter, cleft palate defects occur30. A study tested mutant mice with the CDH1 gene deleted in NCCs by activating Cre recombinase using the Wnt1 promoter (Wnt1-Cre2) and having the Cdh1 gene ‘floxed’ between LoxP sites. The results found that 100% of the mice’s offspring that had both copies deleted had a cleft palate. The craniofacial structures of offspring expressing at least one copy of CDH1 in NCCs developed typically. This shows that for cleft palate to occur, both gene alleles need to be mutated.
The pro-inflammatory environment created by GDM is another factor that can contribute to the development of clefts. Research has shown that after exposing NCC’s to lipopolysaccharide (LPS) + adenosine triphosphate (ATP), both proinflammatory stimuli, a pro-inflammatory response is triggered. The study observed that the LPS + ATP treated cells exhibited reduced dispersion and slower migration speeds. Further analysis revealed that exposure to LPS and ATP CDH1 caused a significant decrease in CDH1 mRNA levels in NCCs, suggesting that proinflammatory signals can downregulate CDH1 expression. Additionally, the study found that the pro-inflammatory environment led to hypermethylation of the CDH1 promoter by about 15%, a change that impairs NCC migration, as seen in the reduced migration distances of treated embryos. These findings indicate that maternal inflammation caused by GDM can influence the microbiome, leading to changes that trigger inflammatory responses. This inflammation, in turn, may lead to CDH1 promoter hypermethylation and decreased CDH1 expression, ultimately disrupting NCC migration and increasing the risk of cleft lip or palate in offspring30.
Conclusion
This paper aimed to further understand the connections between GDM on the maternal gut microbiome and the microbiome’s effects on embryonic cleft palate formation. The gut microbiome is a sensitive, ever-changing system. Studies indicate that hyperglycemia alters the maternal microbiome, decreasing beneficial bacteria while increasing inflammatory microbes. One significant consequence of these microbial changes is an increase in Vitamin A, which may lead to excessive retinoic acid, a known inducer of cleft palate. RA overexpression disrupts essential signaling pathways such as TGFB, SHH, and FGF, which are critical for orofacial development.
Additionally, RA has been shown to influence microbiome composition by altering Lactobacillus levels in pregnant women. Higher levels of Lactobacillus are not only found in females with GDM but also in their cleft-affected babies, supporting the conclusion that excess Lactobacillus may promote cleft defects. Cleft palate arises due to Wnt-signaling dysfunctions, which can result from altercations and irregularities in the maternal microbiome caused by GDM. Dysregulated Wnt signaling impacts craniofacial morphogenesis, contributing to improper palatal fusion. Furthermore, studies have found that specific pathogens, specifically bacterial CW fragments, can cross the placental barrier, potentially influencing fetal development. Once in fetal circulation, bacterial CW triggers inflammation and induces cell death, which can contribute to cleft palate formation. These findings support that maternal inflammatory environments can arise due to increased glucose levels, affecting neural crest cells (NCCs), the precursors to craniofacial structures. In response to inflammation, NCCs alter their migratory behavior and increase the expression of pro-inflammatory cytokines such as TNF-α, IL-1β, IL-6, and NF-κB. Cytokines are inflammatory molecules secreted by immune system cells and can cause the hypermethylation of CDH1, the gene encoding E-cadherin. E-cadherin is related to cleft lip/palate development and is essential for the proper migration of NCC. Studies have found that cleft defects occur when there is a mutation and/or hypermethylation of CDH1.
Furthermore, NCCs exposed to pro-inflammatory stimuli, such as LPS + ATP, show decreased dispersion, reduced migration speed, and significantly lower CDH1 mRNA levels, further linking inflammation to cleft formation. The research behind GDM and cleft palate development is still emerging, and information and resources remain limited. Despite its rising popularity, even research on the gut microbiome is not fully understood. However, as global rates of GDM increase, further research is needed to clarify the relationship between GDM, microbiome alterations, and cleft palate. Understanding these connections can improve prenatal care by identifying high-risk pregnancies and developing targeted interventions. Raising awareness of these links can also help women take preventative measures to reduce risk factors associated with GDM. Future research should focus on strategies to reverse or mitigate microbiome alterations caused by GDM, potentially preventing cleft defects.
The mechanistic model outlined,where GDM‑driven microbiome dysbiosis triggers systemic inflammation, CDH1 hyper‑methylation, and Wnt/RA perturbations that may culminate in cleft palate, relies largely on correlative human data and mechanistic insights from murine or in vitro work. At present, no study has quantified CDH1 methylation in placental or cord‑blood samples from GDM pregnancies, prospectively manipulated the maternal microbiome while monitoring orofacial outcomes, or measured RA/Wnt read‑outs in human neural‑crest‑cell models exposed to GDM serum. Therefore, future research should focus on establishing more direct mechanistic links among GDM, microbiome dysbiosis, inflammatory signaling, and orofacial development. Filling these gaps will be essential to confirm or refute the proposed link between GDM and orofacial clefts. Additionally, studies should investigate whether other forms of diabetes, such as type 1 and type 2, similarly influence the microbiome and fetal development. Exploring the roles of different metabolic disorders, including obesity and hypertension, in shaping the maternal microbiota and affecting fetal orofacial formation could provide further insight into congenital abnormalities. Expanding this research will further understanding, which could lead to improved screening, prevention, and treatment strategies for conditions influenced by microbiome disruptions, ultimately reducing the prevalence of cleft defects and other developmental disorders.
Acknowledgments
I would like to express my sincere gratitude to Kiana Manian for her invaluable support and encouragement throughout the development of this project. I am especially thankful to Dr. Annabelle S. Menzies, whose insights and expertise in biomedical research greatly informed and elevated my understanding of this subject. I also thank the National High School Journal of Science for providing a platform for students to share and celebrate scientific research.
References
- S. Pedroza Matute, S. Iyavoo. Exploring the gut microbiota: Lifestyle choices, disease associations, and personal genomics. Front Nutr. 10, Article 1225120 (2023). [↩] [↩] [↩]
- G. A. Cresci, E. Bawden. Gut microbiome. Nutr Clin Pract. 30, 734–746 (2015). [↩]
- S. Migliarini, G. Pacini, B. Pelosi, G. Lunardi, M. Pasqualetti. Lack of brain serotonin affects postnatal development and serotonergic neuronal circuitry formation. Mol Psychiatry. 18, 1106–1118 (2013). [↩]
- F. D. Ihekweazu, J. Versalovic. Development of the pediatric gut microbiome: Impact on health and disease. Am J Med Sci. 356, 413–423 (2018). [↩]
- E. M. Alfadhli. Gestational diabetes mellitus. Saudi Med J. 36, 399–406 (2015). [↩]
- J. F. Plows, J. L. Stanley, P. N. Baker, C. M. Reynolds, M. H. Vickers. The pathophysiology of gestational diabetes mellitus. Int J Mol Sci. 19, 3342 (2018). [↩] [↩]
- National Institute of Dental and Craniofacial Research. Cleft lip & palate. https://www.nidcr.nih.gov/health-info/cleft-lip-palate (2021). [↩]
- A. Babai, M. Irving. Orofacial clefts: Genetics of cleft lip and palate. Genes. 14, 1603 (2023). [↩]
- P. A. Ysunza, M. C. Pamplona, G. Repetto. Cleft palate, interdisciplinary diagnosis, and treatment. Biomed Res Int. 2015, 1–2 (2015). [↩]
- M. A. Shkoukani, M. Chen, A. Vong. Cleft lip – A comprehensive review. Front Pediatr. 1, Article 53 (2013). [↩]
- K. K. J. L. Schuyler, Hunt. The increasing prevalence of diabetes in pregnancy. https://pmc.ncbi.nlm.nih.gov/articles/PMC2043158 (2008). [↩]
- R. F. Ionescu, R. M. Enache, S. M. Cretoiu, B. S. Gaspar. Gut microbiome changes in gestational diabetes. Int J Mol Sci. 23, 12839 (2022). [↩]
- F. Magne, M. Gotteland, L. Gauthier, A. Zazueta, S. Pesoa, P. Navarrete, R. Balamurugan. The Firmicutes/Bacteroidetes ratio: A relevant marker of gut dysbiosis in obese patients? Nutrients. 12, Article 1474 (n.d.). [↩]
- S. Dias, C. Pheiffer, S. Adam. The maternal microbiome and gestational diabetes mellitus: Cause and effect. Microorganisms. 11, 2217 (2023). [↩] [↩]
- K. J. Walters, Martiny, E. B. Alpha-, beta-, and gamma-diversity of bacteria varies across habitats. https://pmc.ncbi.nlm.nih.gov/articles/PMC7510982 (2020). [↩]
- T. Chen, Y. Qin, M. Chen, Y. Zhang, X. Wang, T. Dong, G. Chen, X. Sun, T. Lu, R. A. White, P. Ye, H. M. Tun, Y. Xia. Gestational diabetes mellitus is associated with the neonatal gut microbiota and metabolome. BMC Med. 19, Article 129 (2021). [↩]
- K. Gorczyca, A. Obuchowska, Ż. Kimber-Trojnar, M. Wierzchowska-Opoka, B. Leszczyńska-Gorzelak. Changes in the gut microbiome and pathologies in pregnancy. Int J Environ Res Public Health. 19, 9961 (2022). [↩]
- Blood borne: Bacterial components in mother’s blood influence fetal development. Inflamm Cell Signal. 3, Article 1421 (2016). [↩]
- G. W. Ray, Q. Zeng, P. Kusi, H. Zhang, T. Shao, T. Yang, Y. Wei, M. Li, X. Che, R. Guo. Genetic and inflammatory factors underlying gestational diabetes mellitus: A review. Front Endocrinol. 15, Article 1399694 (2024). [↩]
- E. Gershater, Y. Liu, B. Xue, M. K. Shin, H. Koo, Z. Zheng, C. Li. Characterizing the microbiota of cleft lip and palate patients: A comprehensive review. Front Cell Infect Microbiol. 13, Article 1159455 (2023). [↩] [↩]
- G. Duester. Retinoic acid synthesis and signaling during early organogenesis. Cell. 134, 921–931 (2008). [↩]
- S. Wang, Z. Cui, H. Yang. Interactions between host and gut microbiota in gestational diabetes mellitus and their impacts on offspring. BMC Microbiol. 24, Article 3255 (2024). [↩]
- J. Yu, Y. Liu, L. Xu. Associations between serum vitamin A levels in early pregnancy and the risk of gestational diabetes mellitus. Diabetes Metab Syndr Obes. 17, 2895–2901 (2024). [↩]
- B. Sun, K. S. Reynolds, M. A. Garland, M. McMahon, S. K. Saha, C. J. Zhou. Epigenetic implications in maternal diabetes and metabolic syndrome‐associated risk of orofacial clefts. Birth Defects Res. 115, 1835–1850 (2023). [↩] [↩] [↩] [↩]
- E. L. Semble, W. C. Wu. Prostaglandins in the gut and their relationship to non-steroidal anti-inflammatory drugs. Trends Pharmacol Sci. 10, 20 (n.d.). [↩]
- A. M. Tyagi, M. Yu, T. M. Darby, C. Vaccaro, J.-Y. Li, J. A. Owens, E. Hsu, J. Adams, M. N. Weitzmann, R. M. Jones, R. Pacifici. The microbial metabolite butyrate stimulates bone formation via T regulatory cell-mediated regulation of WNT10B expression. Immunity. 49, 1116–1131 (n.d.). [↩]
- G. S. Sun, Vasdev, Martin, Gadag, Zhang. Altered calcium homeostasis is correlated with abnormalities of fasting serum glucose, insulin resistance, and beta-cell function in the Newfoundland population. https://pubmed.ncbi.nlm.nih.gov/16249463 (2005). [↩]
- A. E. J. Tuomanen, Loughra. Blood borne: Bacterial components in mother’s blood influence fetal development. https://pmc.ncbi.nlm.nih.gov/articles/PMC5340199 (2017). [↩]
- S. de A. Passinato Gheller, A. M. Borba, A. N. Porto, K. de A. Veiga, A. Aranha. Periodontal findings in children and adolescents with cleft lip and/or palate: A case-control study. https://pubmed.ncbi.nlm.nih.gov/33892839 (2021). [↩]
- N. Alvizi, Nani, Brito, Kobayashi, Passos-Bueno, L. D. L. G. M. R. Mayor. Neural crest E-cadherin loss drives cleft lip/palate by epigenetic modulation via pro-inflammatory gene–environment interaction. Nat Commun. 14, Article 38526 (2023). [↩] [↩]


