
Abstract
Neurodegenerative diseases are characterized by the progressive loss of neurons caused by mechanisms such as oxidative stress, mitochondrial dysfunction, and apoptosis. Ethanol (EtOH) and glutamate are known to be prominent neurotoxicants that induce neuronal damage through these pathways, but their common cytotoxic mechanisms are poorly understood. In this study, we analyzed changes in cell morphology, viability, reactive oxygen species (ROS) production, and apoptosis in SH-SY5Y neuroblastoma cells to evaluate the cytotoxic effects of EtOH and glutamate. RNA sequencing (RNA-seq) revealed distinct changes in gene expression related to oxidative stress and, in particular, hypoxia signaling pathways, suggesting a common toxic mechanism of both neurotoxins. By qPCR analysis, we identified decreased expression of mitochondrial oxidative phosphorylation (OXPHOS)-related genes, such as ATP5F1A and NDUFA9, and increased expression of SOD1 and NRF2 genes associated with antioxidant responses, demonstrating mitochondrial dysfunction and activation of oxidative stress. Furthermore, we applied hyperoxia and vitamin C to assess their potential protective effects and found that ROS levels were significantly reduced, apoptosis was inhibited, and mitochondria-related gene expression was partially restored. These findings demonstrate that EtOH and glutamate induce neurotoxicity through oxidative stress, mitochondrial dysfunction, and hypoxia pathways, and suggest that hyperoxia and vitamin C may have protective effects to mitigate this damage. These findings provide new insights into the molecular mechanisms of neurodegenerative diseases and may contribute to the development of potential therapeutic strategies to promote neurological health and improve quality of life.
Keywords : SH-SY5Y, Ethanol, Glutamate, ROS, Apoptosis, Hypoxia, RNA Sequencing, High Oxygen, Vitamin C, Neuroprotection
Introduction
Neurodegenerative diseases such as Alzheimer’s disease, Parkinson’s disease, and Huntington’s disease are characterized by progressive neuronal loss and cognitive decline, which are closely associated with mechanisms including oxidative stress, mitochondrial dysfunction, and apoptosis1. Various environmental and endogenous factors contribute to neuronal damage, among which ethanol (EtOH) and glutamate are widely recognized neurotoxicants. EtOH is known to disrupt redox homeostasis by inducing oxidative stress, mitochondrial impairment, and apoptotic pathways, while glutamate triggers excitotoxicity through overactivation of glutamate receptors, leading to calcium overload and excessive ROS production2. Although mechanistically distinct in their initial targets, both compounds converge on shared downstream effects, such as oxidative stress and mitochondrial dysfunction, which are key contributors to neuronal injury.
Oxidative stress, defined as an imbalance between reactive oxygen species (ROS) production and antioxidant defenses, plays a central role in the pathology of neurodegeneration3. Mitochondria, the primary energy producers within cells, are highly susceptible to oxidative damage, resulting in impaired oxidative phosphorylation (OXPHOS), ATP depletion, and initiation of apoptosis4. Gene products such as ATP5F1A and NDUFA9 are essential for OXPHOS5, whereas antioxidant response genes such as SOD1 and NRF2 play protective roles by regulating ROS detoxification and redox signaling5.
In this study, we investigated the molecular responses of SH-SY5Y neuroblastoma cells following exposure to EtOH and glutamate. Transcriptomic profiling revealed not only the expected activation of oxidative stress and apoptosis pathways, but also significant enrichment of hypoxia-related gene signatures. This observation suggests that toxicant-induced mitochondrial dysfunction may result in pseudo-hypoxic states due to altered oxygen consumption and impaired intracellular oxygen availability, even under normoxic culture conditions.
To counteract these stressors, we explored the effects of elevated oxygen conditions (30% O₂, defined in literature as moderate hyperoxia in cell culture models) and vitamin C, a well-established antioxidant. Although hyperoxia can, under some conditions, increase ROS production, it may also alleviate intracellular hypoxic stress by enhancing oxygen diffusion. Vitamin C, in contrast, directly scavenges free radicals and promotes antioxidant gene expression. While the independent effects of oxygen modulation and vitamin C have been studied in various systems, their combined influence in the context of dual oxidative and hypoxic stress has not been thoroughly investigated, particularly in neuronal cells exposed to EtOH and glutamate.
Therefore, this study addresses a critical gap by evaluating how hyperoxia and vitamin C, both individually and in combination, modulate cellular outcomes in a dual-stress neurotoxicity model. Rather than making definitive claims about neuroprotection, our goal is to provide mechanistic insights into the cellular responses elicited by these interventions. Findings from this in vitro investigation may inform the design of future in vivo studies and contribute to the broader understanding of potential strategies for mitigating neurodegenerative damage.
Results
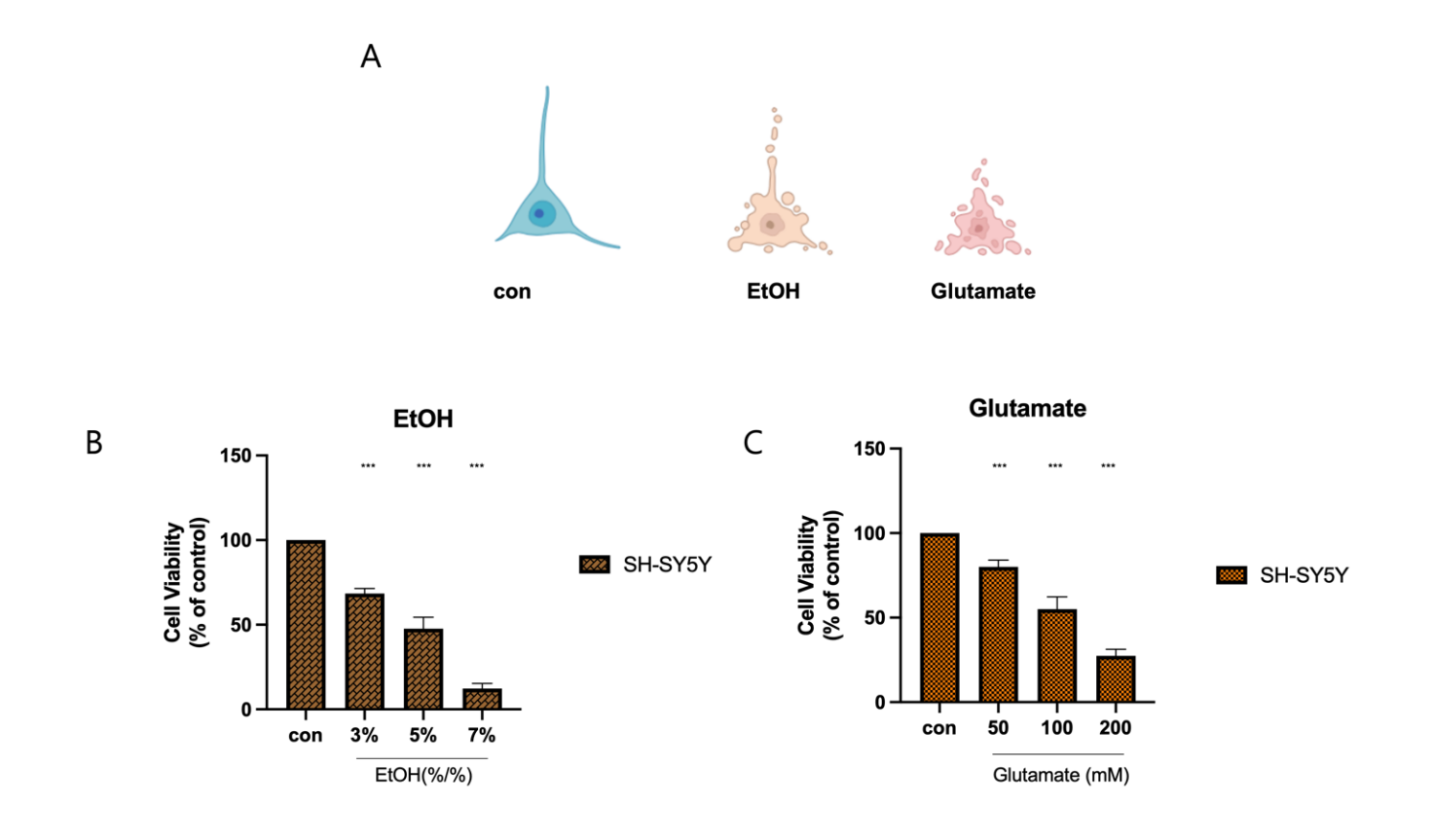
(A) Representative images depicting morphological changes in SH-SY5Y cells after exposure to EtOH and glutamate.
(B) Cell viability assay showing the dose-dependent effect of EtOH on SH-SY5Y cells. Cells were treated with 3%, 5%, and 7% EtOH, with 5% EtOH reducing viability to approximately 50% of the control. Statistical significance is indicated (***p < 0.001)
(C) Cell viability assay assessing the effects of glutamate at concentrations of 50, 100, and 200 mM on SH-SY5Y cells. A significant reduction in viability is observed in a dose-dependent manner, with 100 mM glutamate reducing viability to approximately 50% of the control. Statistical significance is indicated (***p < 0.001)
All data are presented as mean ± SD. Statistical significance was determined using one-way ANOVA followed by Tukey’s post hoc test (*p < 0.05, **p < 0.01, ***p < 0.001). Sample size: n = 3 independent biological replicates.
To determine the cytotoxic effects of ethanol (EtOH) and glutamate on SH-SY5Y cells (Figure 1A), we analyzed cell morphology and viability after increasing concentrations of these neurotoxicants. Morphological examination revealed that control cells maintained an intact neuronal structure, while EtOH-treated cells showed characteristic signs of cellular stress, including shrinkage, and glutamate-treated cells showed severe fragmentation, indicating extensive cell death. Cell viability assays further validated these observations, confirming a dose-dependent decrease in viability. SH-SY5Y cells treated with 3%, 5%, and 7% EtOH showed a progressive decrease in viability, with higher concentrations resulting in near cell death. Notably, approximately 50% cell viability was observed at around 5% EtOH treatment. Similarly, exposure to glutamate at 50, 100, and 200 mM significantly reduced viability, with the highest concentration inducing the most pronounced cytotoxic effect (Figure 1B). In particular, glutamate treatment at approximately 100 mM resulted in about 50% cell viability. Based on these results, subsequent experiments were conducted using 5% EtOH and 100 mM glutamate to achieve consistent cytotoxic conditions. These results confirm that both EtOH and glutamate severely impair SH-SY5Y cell survival through mechanisms involving oxidative stress and cell death, supporting the need for further investigation of potential protective interventions.
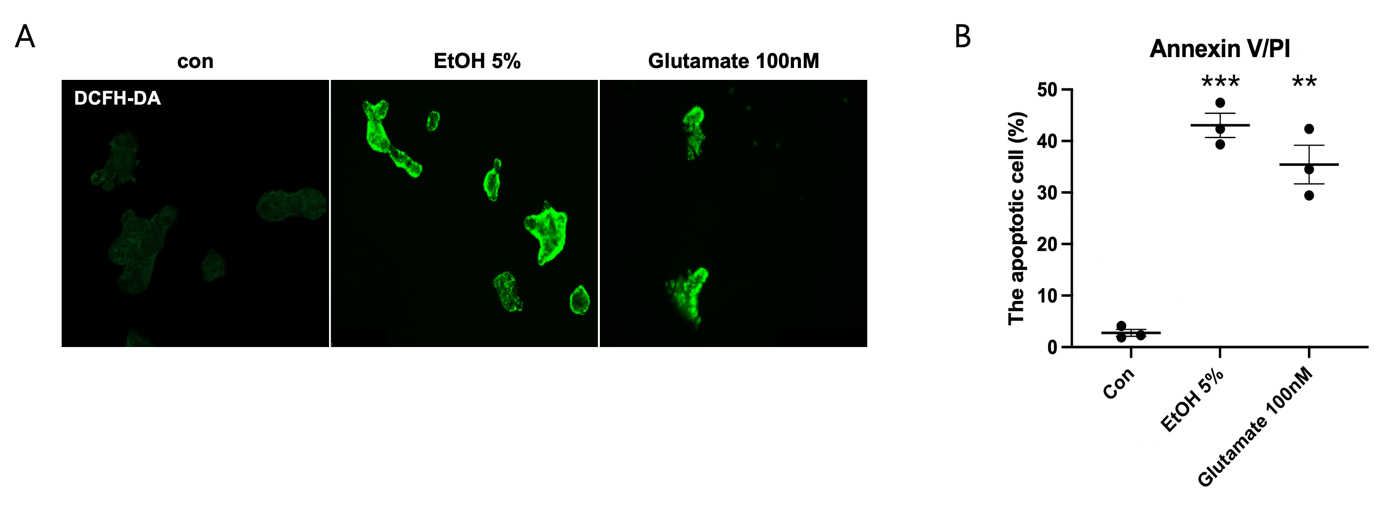
(A) Representative fluorescence images of intracellular reactive oxygen species (ROS) detected using DCFH-DA staining. Control (con) cells exhibit minimal fluorescence, indicating low basal ROS levels. In contrast, SH-SY5Y cells treated with 5% EtOH or 100 mM glutamate show a marked increase in green fluorescence intensity, suggesting elevated ROS production.
(B) Quantification of apoptotic cells using Annexin V/PI staining. The percentage of apoptotic cells significantly increases following exposure to EtOH and glutamate compared to the control group. EtOH treatment results in approximately 40% apoptosis, while glutamate induces a similar apoptotic rate. Statistical significance is indicated (**p < 0.01, ***p < 0.001).
All data are presented as mean ± SD. Statistical significance was determined using one-way ANOVA followed by Tukey’s post hoc test (*p < 0.05, **p < 0.01, ***p < 0.001). Sample size: n = 3 independent biological replicates.
To evaluate the effects of ethanol (EtOH) and glutamate on oxidative stress and apoptosis in SH-SY5Y cells, we assessed ROS production and the percentage of apoptotic cells. Using DCFH-DA staining to visualize intracellular ROS levels, control cells showed little fluorescence, indicating low ROS levels, whereas EtOH (5%) and glutamate (100mM) treatment significantly increased the green fluorescence intensity, suggesting increased ROS production (Figure. 2A). To further confirm the induction of apoptosis, Annexin V/PI staining was performed to quantify the percentage of apoptotic cells. The percentage of apoptotic cells was significantly higher in both EtOH- and glutamate-treated groups compared to the control group, with EtOH exposure inducing approximately 40% apoptosis and glutamate treatment resulting in a similar percentage of apoptosis (Figure. 2B). These results suggest that EtOH and glutamate induce significant oxidative stress and cell death in SH-SY5Y cells, indicating neurotoxicity.
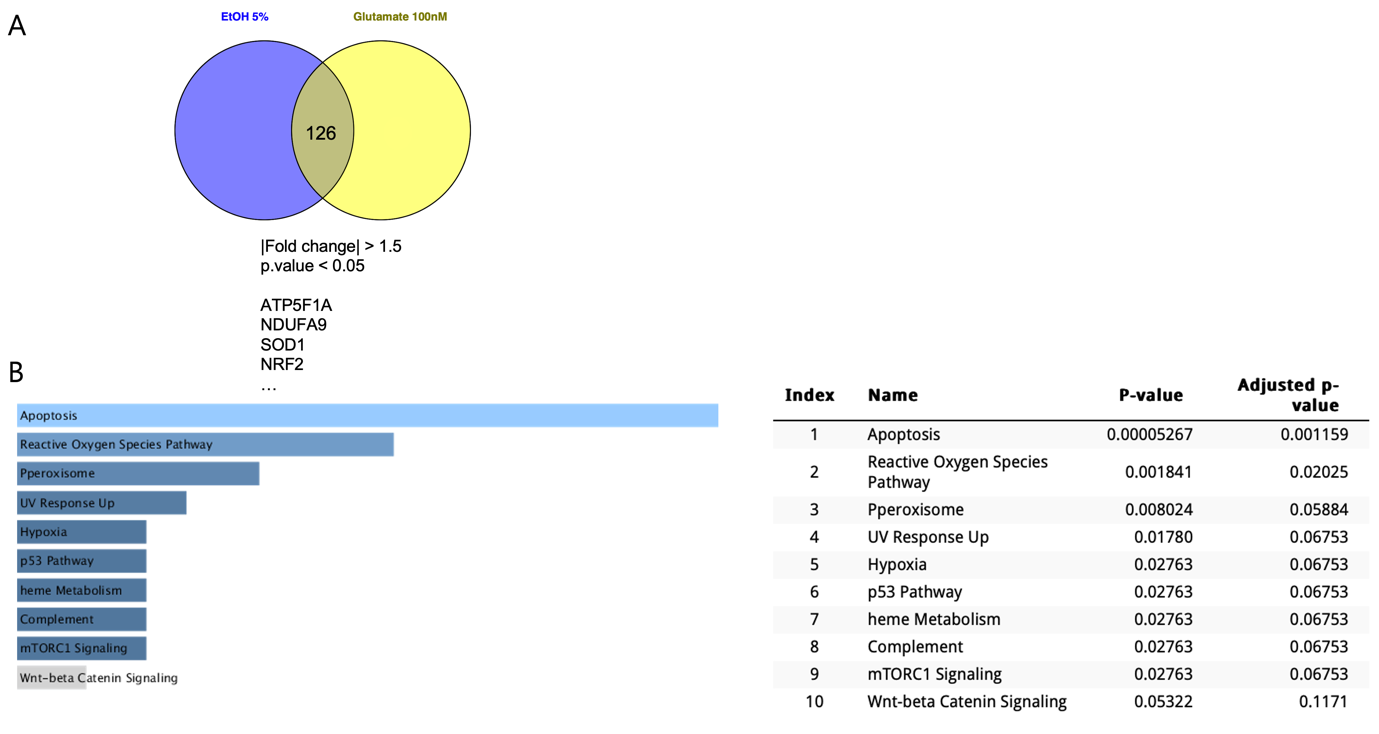
(A) Venn diagram showing 126 commonly altered genes in SH-SY5Y cells treated with 5% EtOH or 100 mM glutamate (|fold change| > 1.5, p < 0.05). Selected differentially expressed genes, including ATP5F1A, NDUFA9, SOD1, and NRF2, are associated with mitochondrial function and oxidative stress regulation.
(B) Pathway enrichment analysis of differentially expressed genes reveals significant upregulation of apoptosis and reactive oxygen species (ROS)-related pathways. Additional pathways related to hypoxia, p53 signaling, and mitochondrial function are also activated, suggesting shared cytotoxic mechanisms. For each pathway, the adjusted p-value (FDR), enrichment score to enrichment are indicated in parentheses. Statistical significance was calculated using Fisher’s exact test followed by Benjamini-Hochberg correction.
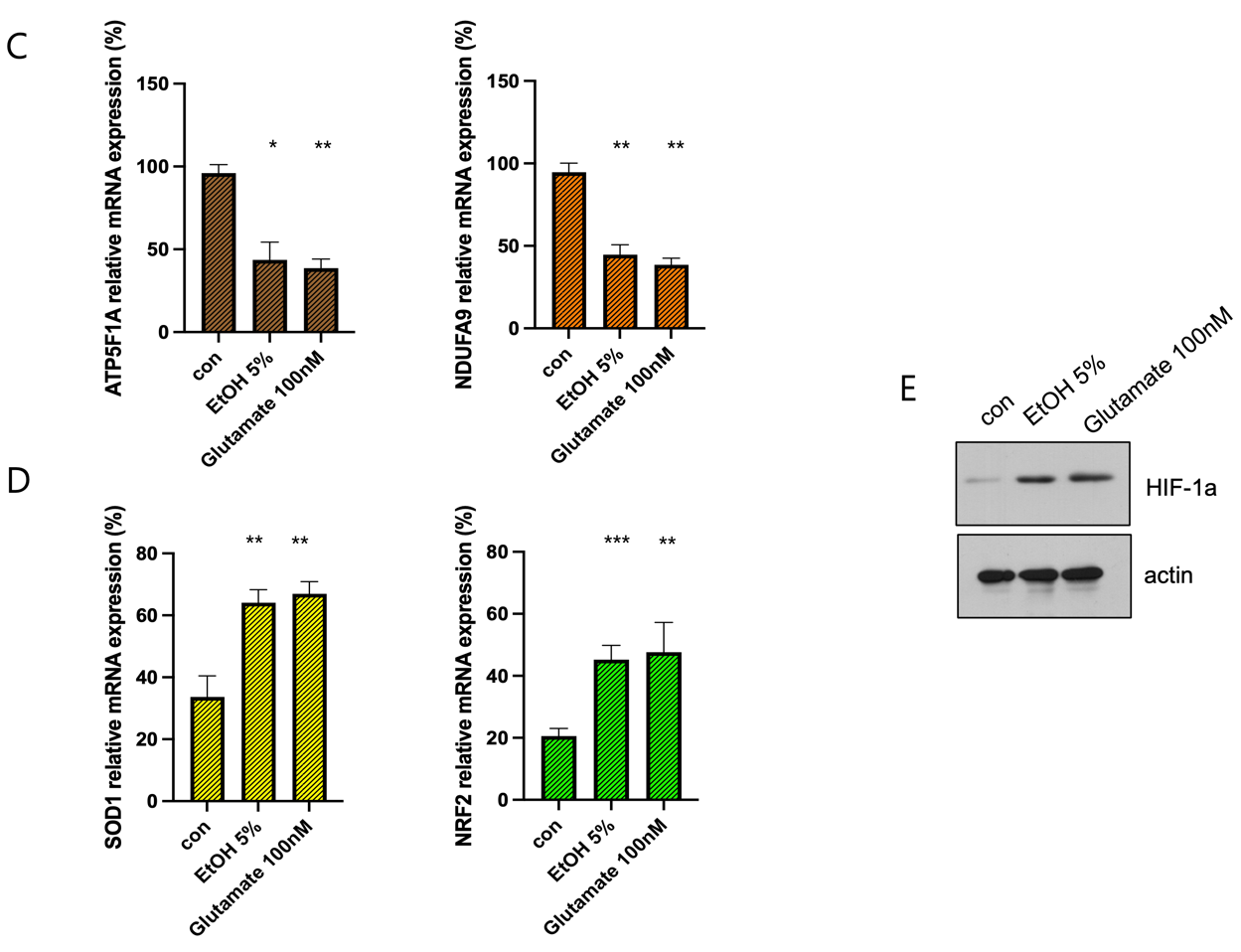
(C) qPCR validation of mitochondrial oxidative phosphorylation (OXPHOS) genes ATP5F1A and NDUFA9. Both genes are significantly downregulated following EtOH and glutamate treatment, indicating mitochondrial dysfunction (**p < 0.01)
(D) qPCR analysis of antioxidant response genes SOD1 and NRF2. Both genes are significantly upregulated in response to EtOH and glutamate exposure, indicating activation of compensatory oxidative stress response mechanisms (***p < 0.001, **p < 0.01).
(E) Western blot analysis of HIF-1a expression under different treatment conditions.
All data are presented as mean ± SD. Statistical significance was determined using one-way ANOVA followed by Tukey’s post hoc test (*p < 0.05, **p < 0.01, ***p < 0.001). Sample size: n = 3 independent biological replicates.
To explore the molecular mechanisms underlying ethanol (EtOH)- and glutamate-induced cytotoxicity in SH-SY5Y cells, we conducted RNA sequencing and gene expression analysis. Comparative analysis identified 126 commonly altered genes in both EtOH- and glutamate-treated groups (|fold change| > 1.5, p < 0.05), suggesting shared cytotoxic mechanisms (Figure. 3A). Pathway enrichment analysis revealed significant upregulation of apoptosis and reactive oxygen species (ROS)-related pathways, along with activation of hypoxia and p53 signaling pathways, with apoptosis and ROS pathways being the most prominent (Figure 3B).
qPCR analysis validated these findings, showing marked downregulation of mitochondrial oxidative phosphorylation (OXPHOS) genes ATP5F1A and NDUFA9 after treatment, indicating mitochondrial dysfunction6 (Figure 3C). Conversely, antioxidant response genes SOD1 and NRF2 were significantly upregulated (Figure 3D), reflecting compensatory response to oxidative stress7. Western blot analysis further confirmed that EtOH and glutamate treatments markedly increased HIF-1α protein levels, supporting the activation of hypoxia signaling pathways under these conditions (Figure 3E).
These results confirm that EtOH and glutamate exposure induces oxidative stress, disrupt mitochondrial function, and trigger cell death in SH-SY5Y cells, with hypoxic and metabolic stress pathways contributing to the cytotoxic effects.
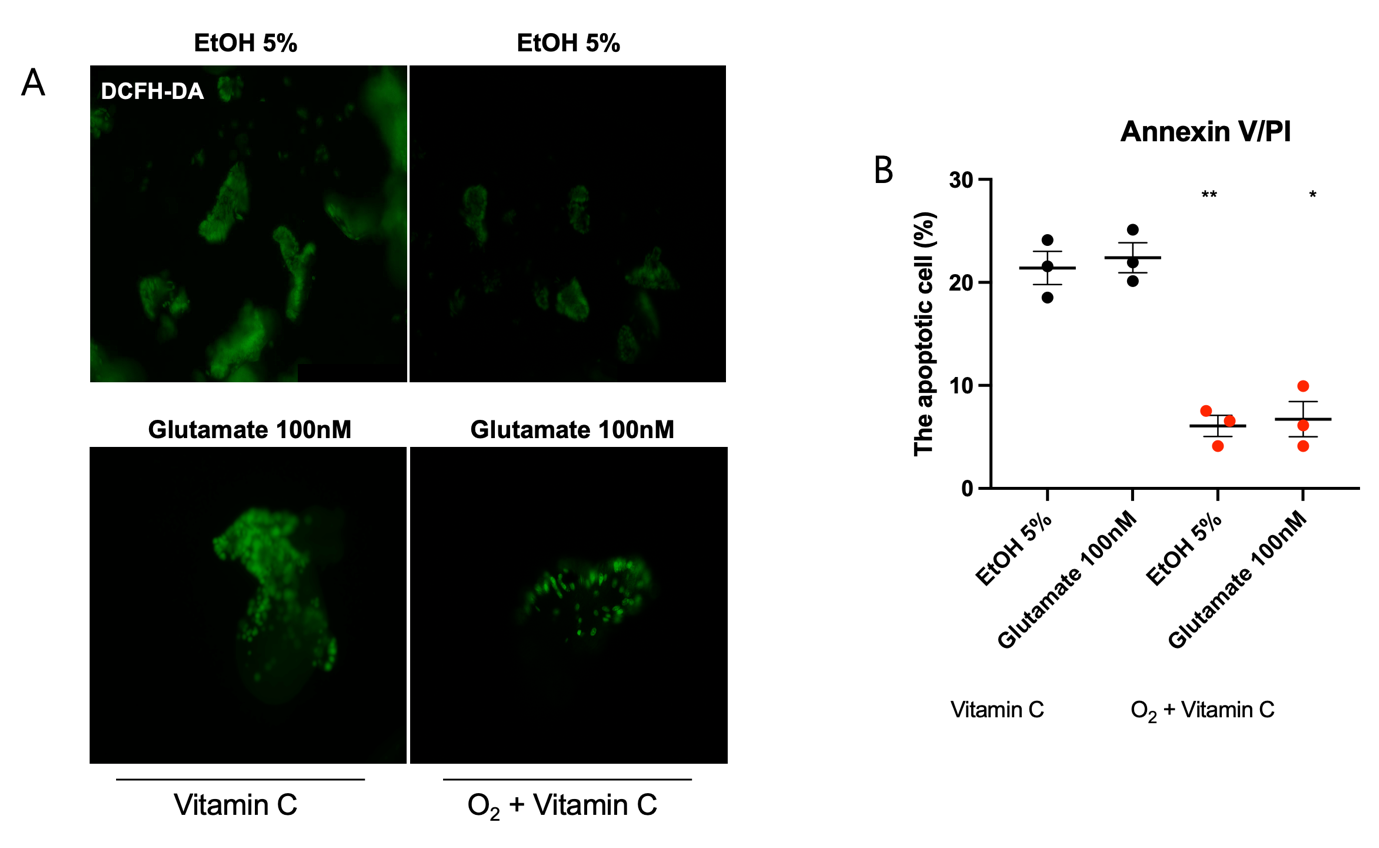
(A) Representative fluorescence images of intracellular reactive oxygen species (ROS) detected using DCFH-DA staining. Treatment with vitamin C alone reduces ROS accumulation, but the combination of hyperoxia and vitamin C results in a more pronounced decrease in fluorescence intensity, indicating a greater reduction in oxidative stress.
(B) Quantification of apoptotic cells using Annexin V/PI staining. Treatment with vitamin C alone decreases apoptosis, while the combination of hyperoxia and vitamin C further reduces apoptotic cell levels, demonstrating an enhanced protective effect (*p< 0.05).
All data are presented as mean ± SD. Statistical significance was determined using one-way ANOVA followed by Tukey’s post hoc test (*p < 0.05, **p < 0.01, ***p < 0.001). Sample size: n = 3 independent biological replicates.
We conducted a study to determine whether increasing the dissolved oxygen concentration in the culture medium helps alleviate hypoxia and whether antioxidant treatment with vitamin C can reduce oxidative stress8. To investigate this, we analyzed the effects of hyperoxia and vitamin C supplementation on ethanol (EtOH)- and glutamate-induced oxidative stress and apoptosis in SH-SY5Y neuroblastoma cells. We hypothesized that adjusting oxygen concentration would mitigate hypoxic stress and that vitamin C, as an antioxidant, would reduce oxidative damage, ultimately preventing neurotoxicity caused by EtOH and glutamate exposure.
To evaluate this, we measured reactive oxygen species (ROS) levels and the percentage of apoptotic cells. Intracellular ROS levels were visualized using DCFH-DA staining. The results showed that in EtOH- and glutamate-treated cells, fluorescence intensity was significantly lower when both hyperoxia and vitamin C were applied together compared to vitamin C treatment alone (Figure 4A), indicating a greater reduction in ROS accumulation.
To further confirm the protective effect against apoptosis, Annexin V/PI staining was performed, and the apoptotic cell population was quantified. Exposure to EtOH and glutamate increased the proportion of apoptotic cells compared to the control group (Figure 2B). However, the combination of hyperoxia and vitamin C significantly reduced the proportion of Annexin V-positive cells compared to vitamin C treatment alone, demonstrating a stronger protective effect against apoptosis (Figure 4B).
These findings suggest that increasing oxygen availability to counteract hypoxia and supplementing antioxidants to combat oxidative stress can effectively mitigate EtOH- and glutamate-induced neurotoxicity.
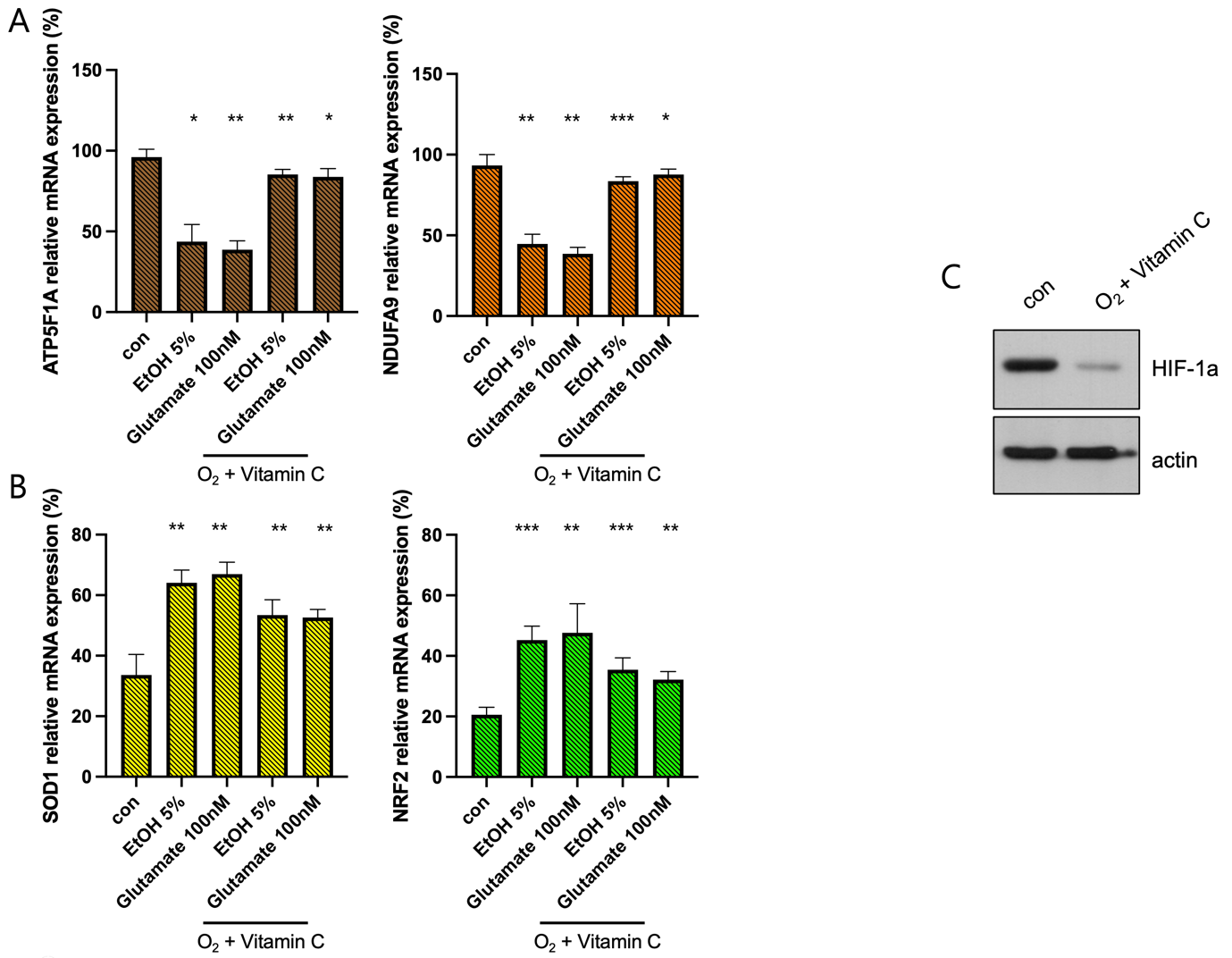
(A) qPCR analysis of mitochondrial oxidative phosphorylation (OXPHOS) genes ATP5F1A and NDUFA9. EtOH (5%) and glutamate (100 mM) exposure significantly downregulate ATP5F1A and NDUFA9 expression, indicating mitochondrial dysfunction. Combined hyperoxia and vitamin C treatment partially restore their expression levels, suggesting improved mitochondrial function (*p < 0.05, **p < 0.01, **p < 0.001).
(B) qPCR analysis of antioxidant response genes SOD1 and NRF2. EtOH and glutamate treatment lead to an upregulation of these genes as a compensatory response to oxidative stress. The combination of hyperoxia and vitamin C reduces this elevation, indicating an overall reduction in oxidative stress burden (**p < 0.01, **p < 0.001).
(C) Western blot analysis of HIF-1a expression under different treatment
All data are presented as mean ± SD. Statistical significance was determined using one-way ANOVA followed by Tukey’s post hoc test (*p < 0.05, **p < 0.01, ***p < 0.001). Sample size: n = 3 independent biological replicates.
To evaluate the effects of hyperbaric oxygen and vitamin C (100 µM) treatment on mitochondrial function and oxidative stress response following ethanol (EtOH) and glutamate exposure, the relative mRNA expression levels of ATP5F1A, NDUFA9, SOD1, and NRF2 were quantified using qPCR. In EtOH- and glutamate-treated cells, combined hyperoxia and vitamin C treatment partially restored the expression of ATP5F1A and NDUFA9, key components of the mitochondrial oxidative phosphorylation (OXPHOS) system, suggesting improved mitochondrial function (Figure. 5A). In addition, SOD1 and NRF2, which are involved in antioxidant defense responses, showed a decrease in elevation (Figure. 5B), indicating a sustained activation of protective mechanisms against oxidative stress. Notably, HIF-1α expression was partially reduced in cells treated with the combination of hyperoxia and vitamin C, suggesting alleviation of hypoxic stress by dual intervention (Figure 3F).
These results suggest that controlled oxygen supplementation and vitamin C administration can effectively mitigate the harmful effects of EtOH and glutamate exposure by preserving mitochondrial integrity and enhancing antioxidant defense pathways in SH-SY5Y cells.
Discussions
1. Cytotoxic Effects of Ethanol and Glutamate
In this study, we investigated the cytotoxicity of ethanol (EtOH) and glutamate in SH-SY5Y neuroblastoma cells. Both compounds induced marked morphological changes, significant reductions in cell viability, and elevated intracellular ROS levels. These effects are consistent with previous reports that EtOH and glutamate trigger neuronal damage through oxidative stress, calcium dysregulation, and mitochondrial impairment9. Although the concentrations used (EtOH 5%, ~855 mM; Glu 100 mM) exceed physiological levels10, they were purposefully selected to establish a robust in vitro model of acute stress. This allowed for the consistent induction of oxidative and apoptotic responses and enabled mechanistic dissection of cellular stress pathways.
2. Molecular Mechanisms and Pathway Activation
ROS quantification revealed substantial oxidative stress following toxicant exposure. While DCFH-DA staining lacks ROS-type specificity and is prone to auto-oxidation, it remains widely used for capturing general oxidative status11. Transcriptomic analysis further demonstrated the upregulation of hypoxia-related genes, which we interpret as pseudo-hypoxic signaling arising from mitochondrial dysfunction and reduced oxygen consumption. These changes may stimulate HIF-1α-driven transcription, a hallmark of metabolic hypoxia in oxidative stress contexts12. To validate RNA-seq results, we analyzed canonical markers of mitochondrial (ATP5F1A, NDUFA9) and antioxidant (SOD1, NRF2) function via qPCR. These genes were chosen based on their well-established roles in OXPHOS and redox regulation5,6,7. Their altered expression reflects both mitochondrial compromise and activation of compensatory antioxidant defenses. Moreover, NRF2 signaling—along with the potential involvement of MAPK cascades—may mediate cellular adaptation to oxidative and hypoxic insults, although pathway-level validation remains a subject for future studies.
3. Protective Effects of Hyperoxia and Vitamin C
We evaluated the ability of hyperoxia and vitamin C, separately and in combination, to mitigate ethanol- and glutamate-induced damage. Hyperoxia (30% O₂) may relieve pseudo-hypoxic stress by increasing oxygen availability, while vitamin C functions as a potent ROS scavenger. Our findings show that combined treatment significantly decreased ROS levels, apoptosis, and partially restored mitochondrial gene expression. These effects likely occur through parallel modulation of mitochondrial respiration and antioxidant signaling, possibly through the mitochondrial-Nrf2-hypoxia axis. While promising, further work is needed to explore whether these interventions influence additional survival or stress response pathways.
4. Limitations and Future Directions
Several limitations must be considered. First, we did not directly quantify dissolved oxygen concentrations or mitochondrial respiration rates, which limits our interpretation of oxygen-driven effects. Second, the study was conducted exclusively in SH-SY5Y cells cultured in 2D conditions, which do not fully recapitulate the complexity of in vivo neural environments. Third, although HIF-1α expression was evaluated, broader profiling of hypoxia markers and pathway components would strengthen the hypoxia-related conclusions. Finally, without in vivo validation, the translational relevance of these findings remains speculative.
In conclusion, our results highlight the converging roles of oxidative stress, mitochondrial dysfunction, and hypoxia-like responses in EtOH- and glutamate-induced neurotoxicity. The combination of hyperoxia and vitamin C offers preliminary evidence of stress mitigation in vitro. Future research using physiologically relevant models and refined oxygen control systems is essential to determine the therapeutic potential of such interventions in neurodegenerative contexts.
Materials and Methods
Cell Culture
SH-SY5Y human neuroblastoma cells were obtained from Korean Cell Line Bank and cultured in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 1% penicillin-streptomycin, and 1% non-essential amino acids. The cells were maintained at 37°C in a humidified atmosphere containing 5% CO₂.
Ethanol and Glutamate Treatment
To induce cytotoxicity, SH-SY5Y cells were treated with varying concentrations of ethanol (3%, 5%, 7%) and glutamate (50 mM, 100 mM, 200 mM) for 24 hours. Based on preliminary results, 5% ethanol and 100 mM glutamate were used for subsequent experiments to achieve consistent cytotoxic conditions.
Hyperoxia and Vitamin C Supplementation
For hyperoxia treatment, cells were cultured in media with increased dissolved oxygen concentrations using a controlled gas incubator set at 30% oxygen. Vitamin C (ascorbic acid, Sigma-Aldrich) was dissolved in sterile PBS to a stock concentration of 10 mM and filtered through a 0.22 µm syringe filter for sterility. To prevent degradation due to oxidation, fresh working solutions (100 µM final concentration) were prepared immediately before each experiment and used within 30 minutes of dilution under light-protected conditions. Cells were treated with hyperoxia, vitamin C, or a combination of both following ethanol and glutamate exposure.
Cell Viability Assay
Cell viability was assessed using the MTT assay. Briefly, MTT solution (0.5 mg/mL) was added to each well and incubated for 4 hours at 37°C. Formazan crystals were dissolved in DMSO, and absorbance was measured at 570 nm using a microplate reader.
Reactive Oxygen Species (ROS) Measurement
Intracellular ROS levels were measured using DCFH-DA staining. Cells were incubated with 10 µM DCFH-DA for 30 minutes at 37°C, washed with PBS, and fluorescence was detected using a fluorescence microscope and quantified with a fluorescence plate reader (excitation/emission: 488/525 nm).
Apoptosis Assay
Apoptotic cells were quantified using Annexin V-FITC/PI double staining. Treated cells were harvested, washed with PBS, and resuspended in binding buffer containing Annexin V-FITC and PI. After incubation for 15 minutes at room temperature in the dark, samples were analyzed by flow cytometry. For accurate quantification of apoptotic populations, gating thresholds were established using unstained and single-stained controls for Annexin V and PI. Compensation controls were applied to adjust for spectral overlap, and standard quadrant separation was used to distinguish viable, early apoptotic, late apoptotic, and necrotic cells.
Protein extraction and SDS-PAGE
SH-SY5Y cells were lysed in RIPA buffer (Thermo Fisher Scientific) supplemented with protease and phosphatase inhibitor cocktails (Sigma-Aldrich) on ice for 30 minutes. Lysates were centrifuged at 14,000 × g for 15 minutes at 4°C, and the supernatant was collected. Protein concentration was determined using the BCA assay (Pierce™ BCA Protein Assay Kit, Thermo Fisher). Equal amounts of protein (20–30 µg) were loaded onto a 10% SDS-PAGE gel, separated by electrophoresis, and transferred onto PVDF membranes (Millipore).
Antibodies
Membranes were blocked with 5% non-fat milk in TBST for 1 hour at room temperature, followed by overnight incubation at 4°C with the following primary antibodies:
Anti-HIF-1α (rabbit monoclonal, 1:1000, Cell Signaling Technology, #36169), Anti-β-actin (mouse monoclonal, 1:5000, Sigma-Aldrich, A5441)
RNA Extraction and qPCR Analysis
Total RNA was extracted using Trizol reagent (Invitrogen) following the manufacturer’s instructions. cDNA was synthesized from 1 µg of total RNA using High Capacity cDNA Reverse Transcription kit (Takara). Quantitative PCR (qPCR) was performed using SYBR Green Master Mix on a Real-Time PCR Detection machine (Bio-rad). Relative gene expression levels of ATP5F1A, NDUFA9, SOD1, and NRF2 were normalized to GAPDH and calculated using the ΔΔCt method. ATP5F1A Forward : CAGTATCGTGAGGTTGCTGC, Reverse : TGTAATCTTGCTGGGCTCCA; NDUFA9 Forward : GATGCATTTCCGGAAGCCAT, Reverse : TGGGACCAACGAAAGCAAAG; SOD1 Forward : GGAGACTTGGGCAATGTGAC, Reverse : CACAAGCCAAACGACTTCCA; NRF2 Forward : GGTTGCCCACATTCCCAAAT, Reverse : AGCAATGAAGACTGGGCTCT
RNA Sequencing and Data Analysis
RNA-seq libraries were prepared from total RNA using Illumina TruSeq Stranded Library Kit (and sequenced on an Illumina NovaSeq 6000. Differential gene expression analysis was performed using DESeq2 (Bioconductor), with |fold change| > 1.5 and p < 0.05 considered significant. Pathway enrichment analysis was conducted using Enrich R.
Statistical Analysis
Data are presented as mean ± standard deviation (SD) from at least three independent experiments. Statistical significance was determined using one-way ANOVA. A p-value < 0.05 was considered statistically significant.
References
- Wimo A., Guerchet M., Ali G.C., Wu Y.T., Prina A.M., Winblad B., Jonsson L., Liu Z., Prince M. The worldwide costs of dementia 2015 and comparisons with 2010. Alzheimers Dement. 13:1–7 (2017), Ferri CP, Prince M, Brayne C, Brodaty H, Fratiglioni L, Ganguli M, Hall K, Hasegawa K, Hendrie H, Huang Y, Jorm A, Mathers C, Menezes PR, Rimmer E, Scazufca M. Global prevalence of dementia: A Delphi consensus study. Lancet. 366:2112–2117 (2005). [↩]
- Yang, F., Luo, J. Endoplasmic Reticulum Stress and Ethanol Neurotoxicity. Biomolecules. 5, 2538-2553 (2015). [↩]
- Mei Ling Jin, Sun Young Park, Young Hun Kim, Jeong-Il Oh, Sang Joon Lee, Geuntae Park, The neuroprotective effects of cordycepin inhibit glutamate-induced oxidative and ER stress-associated apoptosis in hippocampal HT22 cells, NeuroToxicology. 41, 102-111 (2014). [↩]
- Wimo A., Guerchet M., Ali G.C., Wu Y.T., Prina A.M., Winblad B., Jonsson L., Liu Z., Prince M. The worldwide costs of dementia 2015 and comparisons with 2010. Alzheimers Dement. 13:1–7 (2017). [↩]
- Zayed, M., Jeong, BH. Adipose-Derived Mesenchymal Stem Cell Secretome Attenuates Prion Protein Peptide (106–126)-Induced Oxidative Stress via Nrf2 Activation. Stem Cell Rev and Rep (2024), Hammad M, Raftari M, Cesário R, Salma R, Godoy P, Emami SN, Haghdoost S. Roles of Oxidative Stress and Nrf2 Signaling in Pathogenic and Non-Pathogenic Cells: A Possible General Mechanism of Resistance to Therapy. Antioxidants (Basel), 12, no. 7 (2023). [↩]
- Piñero-Pérez R, López-Cabrera A, Álvarez-Córdoba M, Cilleros-Holgado P, Talaverón-Rey M, Suárez-Carrillo A, Munuera-Cabeza M, Gómez-Fernández D, Reche-López D, Romero-González A, Romero-Domínguez JM, de Pablos RM, Sánchez-Alcázar JA. Actin Polymerization Defects Induce Mitochondrial Dysfunction in Cellular Models of Nemaline Myopathies. Antioxidants. 12(12) (2023). [↩]
- Zayed, M., Jeong, BH. Adipose-Derived Mesenchymal Stem Cell Secretome Attenuates Prion Protein Peptide (106–126)-Induced Oxidative Stress via Nrf2 Activation. Stem Cell Rev and Rep (2024). [↩]
- Ruano, C.S.M., Miralles, F., Méhats, C., Vaiman, D. The Impact of Oxidative Stress of Environmental Origin on the Onset of Placental Diseases. Antioxidants. 11, 106 (2022). [↩]
- Antonio González, José A. Pariente, Ginés M. Salido. Ethanol stimulates ROS generation by mitochondria through Ca2+ mobilization and increases GFAP content in rat hippocampal astrocytes. Brain Research, 1178, 28-37 (2007). [↩]
- Donejko M, Rysiak E, Galicka E, Terlikowski R, Głażewska EK, Przylipiak A. Protective Influence of Hyaluronic Acid on Focal Adhesion Kinase Activity in Human Skin Fibroblasts Exposed to Ethanol. Drug Des Devel Ther vol. 11, 2017, pp. 669-76 (2017). [↩]
- Murphy MP, Bayir H, Belousov V, Chang CJ, Davies KJA, Davies MJ, Dick TP, Finkel T, Forman HJ, Janssen-Heininger Y, Gems D, Kagan VE, Kalyanaraman B, Larsson NG, Milne GL, Nyström T, Poulsen HE, Radi R, Van Remmen H, Schumacker PT, Thornalley PJ, Toyokuni S, Winterbourn CC, Yin H, Halliwell B. Guidelines for Measuring Reactive Oxygen Species and Oxidative Damage in Cells and in Vivo. Nat Metab. vol. 4, no. 6, pp. 651-62 (2022). [↩]
- Zhang J, Gao P, Chang WR, Song JY, An FY, Wang YJ, Xiao ZP, Jin H, Zhang XH, Yan CL. The Role of Hif-1alpha in Hypoxic Metabolic Reprogramming in Osteoarthritis. Pharmacol Res, vol. 213, 2025, p. 107649 (2025). [↩]


