
Abstract
Gastric cancer is the fourth most common malignant disease and poses a significant health risk to many. To address this, researchers are currently investigating different types of cell death beyond the well-studied apoptosis that is strongly correlated with tumor evolution. One of the emerging topics of interest is the newly discovered inflammatory pyroptosis cell death pathway, which may be relevant in cancers correlated with bacterial infection, such as gastric cancer. This study aims to provide gastric cancer profiles of the pyroptosis-related cell death gene set for potential prognostic and diagnostic purposes, as well as future potential therapeutic targets for cancer in a clinical setting. Multi-omic data for 21 target genes involved in pyroptosis were acquired and utilized: CASP1, CASP4, CASP5, GSDMD, NLRP1, NLRP3, NLRC4, AIM2, ELANE, PYCARD, GBP1, GBP2, APIP, GZMA, GZMB, NAT10, TRIM21, DPP8, DPP9, MIR223, and ZEB2. By analyzing over 450 gastric cancer tissue samples, comprehensive genetic profiles of mRNA expression and correlating cancer survival were created. Findings of this study revealed that the differential mRNA expression was noteworthy: GBP1, NAT10, DPP9, NLRC4, GZMB, and AIM2 were all upregulated, while ZEB2 and ELANE were downregulated. Further analysis in stomach adenocarcinoma samples showed that overexpression of ZEB2, NLRP1, GBP2, and ELANE is associated with a higher risk of patient mortality compared to other genes, with ZEB2 being the most statistically significant. This indicates the potential role of ZEB2 and ELANE as key biomarkers in pyroptosis cell death. These results indicate that pyroptosis might be involved in cancer development and is potentially useful for cancer diagnosis and prognosis. These singular cancer genetic profiles offer useful information and overall better understanding of pyroptosis and its dual activity in gastric cancer.
Keywords: gastric cancer, biomarker, mRNA expression, non-apoptotic cell death, pyroptosis
Introduction
Following the COVID-19 pandemic, cancer has become an increasingly urgent public health concern with more than 2,001,140 new cancer cases and 611,720 cancer deaths projected to occur in the United States alone1. Contributing to this concern is the upsurge in increasing incidence for 6 of the top 10 cancers (e.g. gastric, breast, uterine, pancreas)2. As the fourth most common malignant disease and the second leading cause of cancer-related death worldwide, gastric cancer (GC) is one of the most prevalent malignant and aggressive cancer types with a 5-year survival3. With a poorly understood carcinogenesis at molecular levels, GC is a heterogeneous disease with various molecular subtypes of significant differing clinical implications, thus demanding extensive research to enhance early detection and treatment strategies4. Biomarkers are characteristics that are objectively measured and evaluated as an indicator of normal biological processes, pathogenic processes, or pharmacological responses to a therapeutic intervention5. Identifying biomarkers through clinical studies and genomic analyses has the potential to significantly enhance patient diagnosis and prognosis, thereby advancing therapeutic strategies for gastric cancer. These efforts could reduce gastric cancer-related mortality rates and improve the chances of successful treatment for a larger number of patients.
The rapid advancement in studies of programmed cell death has opened new possibilities for its application in gastric cancer diagnosis, prognosis, and therapy. An expedited number of studies have highlighted non-apoptotic regulated cell death subroutines and their roles in tumorigenesis such as necroptosis, ferroptosis, entosis, and lysosome-dependent cell death6. A recent study conducted by Jia et al. (2023) has revealed pyroptosis as a newly discovered form of programmed cell death triggered by the activation of inflammasomes7. The paper describes latest advancements on the mechanisms of the pyroptosis pathway, mediated by critical inflammatory caspases7. Pyroptosis, as a form of cellular host defense, plays an important role during bacterial infection where it can promote an inflammatory response and thus eliminate the replicative niche of intracellular pathogens8. Pyroptosis is executed by protein gasdermin D (GSDMD) that is cleaved and activated to release its N-terminal domain by caspase-1/11/4/59. The N terminus of GSDMD subsequently binds membrane liquids and forms holes in the cell membrane, manifesting as changes in cell osmotic pressure and gradual expansion until the cytomembrane ruptures and releases cellular contents with strong inflammatory response mechanisms10. This cell death is distinct from apoptosis as it involves caspase-1, which plays a pivotal role in bacteria-induced cell death not observed in other pathways7. Currently, the definition of pyroptosis encompasses cell death triggered by caspases beyond caspase-17.
Inducing pyroptosis in cancer cells has been suggested as a new target for cancer treatment. However, researchers have shown that pyroptosis may play dual roles in tumor activity. Pyroptosis potentially provides a chronic inflammatory environment aiding tumor cell transition and immune escape, while appropriate levels of pyroptosis have also been shown to enhance the antitumor efficacy of immune checkpoint inhibitors7. Tumorigenesis is regulated by various factors such as the immune microenvironment and chronic inflammation9. In pyroptosis, the released cytokines and growth factors, such as IL-1 and IL-18, play a key role in promoting tumor infiltration and progression. These cytokines stimulate cell proliferation and reduce cell death by activating transcription factors like NF-κB, STAT3, and AP-1 in premalignant cells11. This activation induces genes that promote cell growth and survival11. Pyroptosis, as part of inflammation-driven tumor promotion, may also involve the angiogenic switch, which enables the blood supply necessary for the continued growth of previously dormant tumors11. On the contrary, recent studies also suggest that pyroptosis can exert antitumor effects by altering the tumor microenvironment, which triggers an influx of lymphocytes. This immune response creates a cycle of immune-driven tumor cell destruction, enhancing the vulnerability of tumor cells that were previously resistant to immune checkpoint inhibitors, thereby improving their responsiveness to treatment12.
In the context of GC, over 90% of gastric cancer cases are strongly correlated to H.pylori infection, a common type of bacterial infection that infects around 2 in 3 adults worldwide, which makes it the strongest risk factor for GC13. Due to the role of pyroptosis in combating bacterial infections, such as H.pylori, that are significantly related to incidences of gastric cancer using inflammatory responses, it is critical to explore the unclear potential impacts of pyroptosis on the clinical prognosis and diagnosis of GC. Since a majority of GC cases are H.pylori-associated, examining the activity of the pyroptosis pathway commonly used to combat H.pylori may be a strong indicator of gastric cancer progression and response to treatment. Especially with the current lack of literature exploring the intersection of pyroptosis and GC in general, this study holds extreme prevalence. To do so, the researcher analyzed expression and survival data of key genes in the pyroptosis pathway in GC to observe any correlation indicating pyroptosis as a biomarker of GC.
This study focused on 21 total genes involved in the pyroptosis inflammatory pathway including CASP1, CASP4, CASP5, GSDMD, NLRP1, NLRP3, NLRC4, AIM2, ELANE, PYCARD, GBP1, GBP2, APIP, GZMA, GZMB, NAT10, TRIM21, DPP8, DPP9, MIR223, and
ZEB2. Some of these genes, such as GZMA, GZMB, DPP8, and DPP9 play important roles in positively regulating and inducing pyroptosis. Other genes like APIP negatively regulate pyroptosis through caspase-1-binding inhibition and are known to promote tumorigenesis14. Many genes were selected for this study because they played highly active roles in pyroptosis pathway activation or deactivation. The specific functions of these genes in pyroptosis is listed in Table 1. Understanding the role of these genes in gastric cancer development is crucial to comprehending the complexities of gastric cancer with respect to induced pyroptosis.
This study aimed to provide a comprehensive genomics and clinical association profiling of the 21 pyroptosis cell death genes for future references. The 21 pyroptosis cell death genes included in this study have important functions in the stages of pyroptosis and insight into these genes in GC is necessary to understand possible pyroptosis-related tumorigenesis or tumor prevention. With the growing focus on pyroptosis in cancer research, these timely profiles will offer valuable genetic insights and serve as a helpful resource for future studies on the role and potential of pyroptosis-related cell death as a cancer biomarker in gastric cancer, which could revolutionize GC treatment and development in clinical settings.
| Gene Symbol | Full name | Function in Pyroptosis |
| CASP1 | Caspase 1 | Cleaves pro-interleukin-1beta to play an immunomodulatory function15 |
| CASP4 | Caspase 4 | Cleaves Gasdermin-D to release its N-terminal domain16 |
| CASP5 | Caspase 5 | Cleaves Gasdermin-D to release its N-terminal domain16 |
| GSDMD | Gasdermin-D | Binds membrane lipids which leads to change |
| in osmotic pressure and swelling16 | ||
| NLRP1 | NLR Family PyrinDomain Containing 1 | Mediates inflammasome activation in response to various pathogen-associated signals17 |
| NLRP3 | NLR Family PyrinDomain Containing 3 | Mediates inflammasome activation in response to defects in membrane integrity, leading tosecretion of inflammatory cytokines IL1B and IL1818 |
| NLRC4 | NLR CARD Domain Containing 4 | Pathogenic bacteria protein sensor which responds by assembling an inflammasome complex19 |
| AIM2 | Absent in Melanoma 2 | Sensor component of the AIM2 inflammasome20 |
| ELANE | Elastase, Neutrophil Expressed | GSDMB cleavage promoter and pyroptosis inhibitor21 |
| PYCARD | PYD and CARDDomain Containing | Caspase-mediated cell death promoter22 |
| GBP1 | Guanylate Binding Protein 1 | Interferon (IFN)-inducible GTPase that is a positive regulator of inflammasome assembly23 |
| GBP2 | Guanylate Binding Protein 2 | Interferon (IFN)-inducible GTPase that is a positive regulator of inflammasome assembly24 |
| APIP | APAF1 Interacting Protein | CASP1-dependent pyroptosis inhibitor25 |
| GZMA | Granzyme A | Protease in the cytosolic granules of cytotoxic T-cells and NK-cells that activatescaspase-independent pyroptosis26 |
| GZMB | Granzyme B | Protease in the cytosolic granules of cytotoxic T-cells and NK-cells that activatescaspase-independent pyroptosis27 |
| NAT10 | N-Acetyltransferase 10 | Negative regulator of neutrophil pyroptosis28 |
| TRIM21 | Tripartite Motif Containing 21 | Pyroptosis promoter through Lys6′-linked ubiquitination of ISG12a/IFI2729 |
| DPP8 | Dipeptidyl Peptidase 8 | Key inhibitor of caspase-1 monocyte and macrophage pyroptosis30 |
| DPP9 | Dipeptidyl Peptidase 9 | Key inhibitor of caspase-1 monocyte and macrophage pyroptosis30 |
| MIR223 | MicroRNA 223 | Up-regulates expression of caspase-131 |
| ZEB2 | Zinc Finger E-BoxBinding Homeobox 2 | Knockdown of ZEB2 induces pyroptosis32 |
Methods
Data Acquisitions
The data for RSEM-normalized mRNA expression and clinical survival were collected from The Cancer Genome Atlas database (TCGA). The mRNA expression data has been downloaded from UCSC Xena, which corrects for varying batch effects across TCGA cohort samples. From this database, 450 samples of stomach adenocarcinoma data, with 32 paired tumor and normal samples, were used for this study for further analysis using the Gene Set
Cancer Analysis tool (GSCA)33.
Expression Analysis
A differential analysis was conducted using the GSCA tool to compare expression levels between tumor and normal samples, alongside an examination of differential expression for individual genes. All expression analyses, along with the plotting, were carried out using R Foundation for Statistical Computing (2020), version 4.0.3, and ggplot2 (v3.3.2), or obtained from the corresponding data source websites34. The significance of differences between the two groups were evaluated using the Wilcoxon rank sum test. For data comparison, the integrated
expression level of the pyroptosis gene set was calculated using the Gene Set Variation Analysis (GSVA) with the R package GSVA35.
Survival Analysis
The mRNA expression levels and clinical survival data were assessed using GSCA,
categorizing tumor samples into high and low-expression groups based on the median gene value provided by RSEM, a software tool that quantifies transcript abundances from RNA-Seq data
and aids with the differential expression analyses. RSEM normalized scores correct for various biases in RNA-Seq data, such as sequencing depth and the length of transcripts, to provide
comparable expression levels across differing samples. The survival analysis was conducted using the R package, fitting survival time and status between the groups. The Cox Proportional-Hazards model calculated the survival risk (Hazard Ratio, HR) for each gene, and log-rank tests were performed using the Kaplan-Meier method for individual genes in stomach adenocarcinoma36).
Results
GC Gene Expression Analysis Profiles of Pyroptosis Gene Set
The expression difference of pyroptosis genes between cancer and non-cancer tissues from TCGA was analyzed. The analysis of the expression profiles of pyroptosis cell death genes in cancer reveal differential expression, though the degree of overexpression or underexpression varied. The findings indicated GBP1, NAT10, DPP9, NLRC4, GZMB, and AIM2 all appeared to be up-regulated while ZEB2 and ELANE appeared to be down-regulated (Figure 1). Out of all the genes within the gene-set, ELANE has the most differential average expression between tumor and normal samples (Figure 2). However, the expression of these genes showed significant differences when comparing cancerous and non-cancerous tissues.
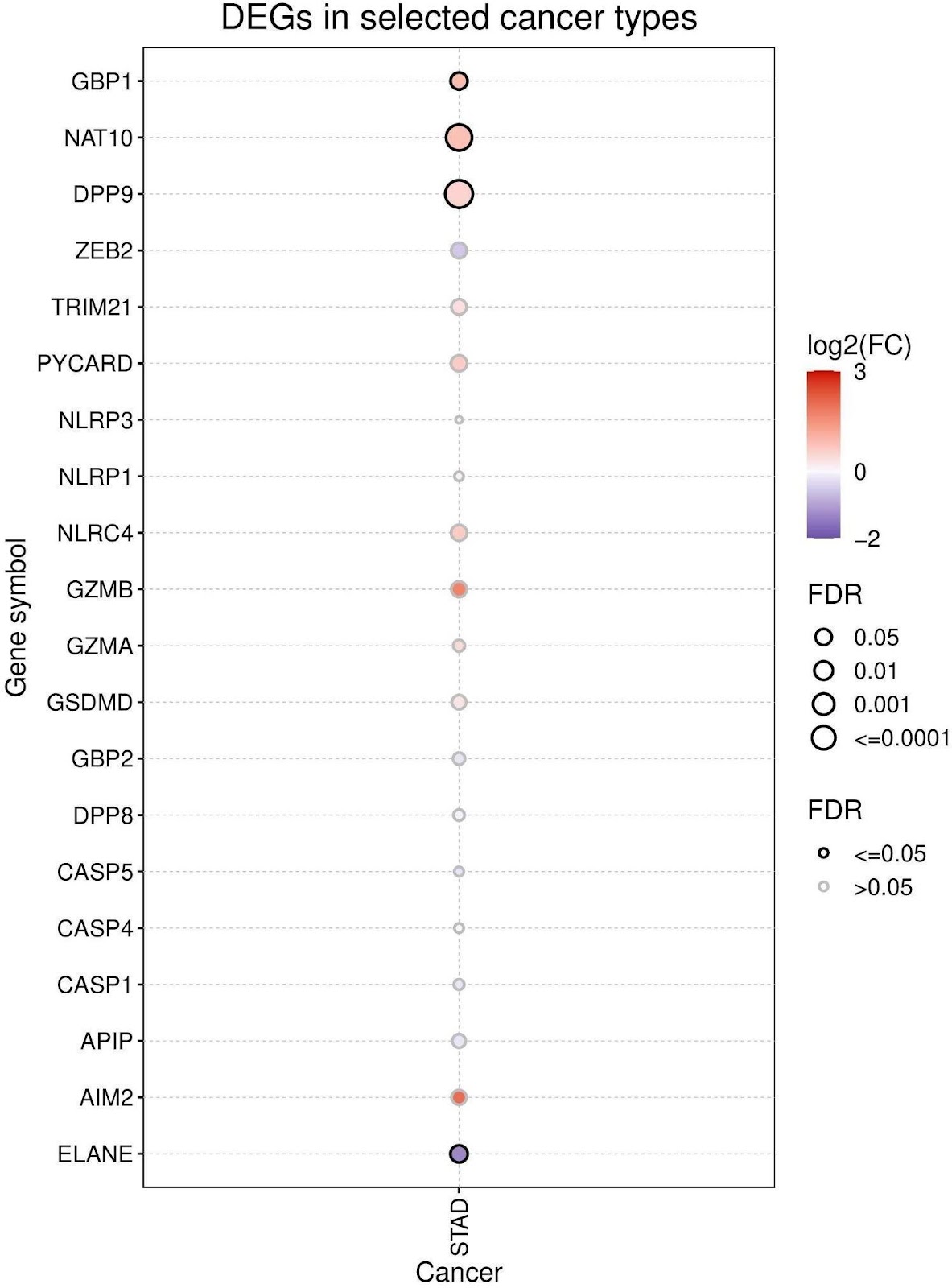
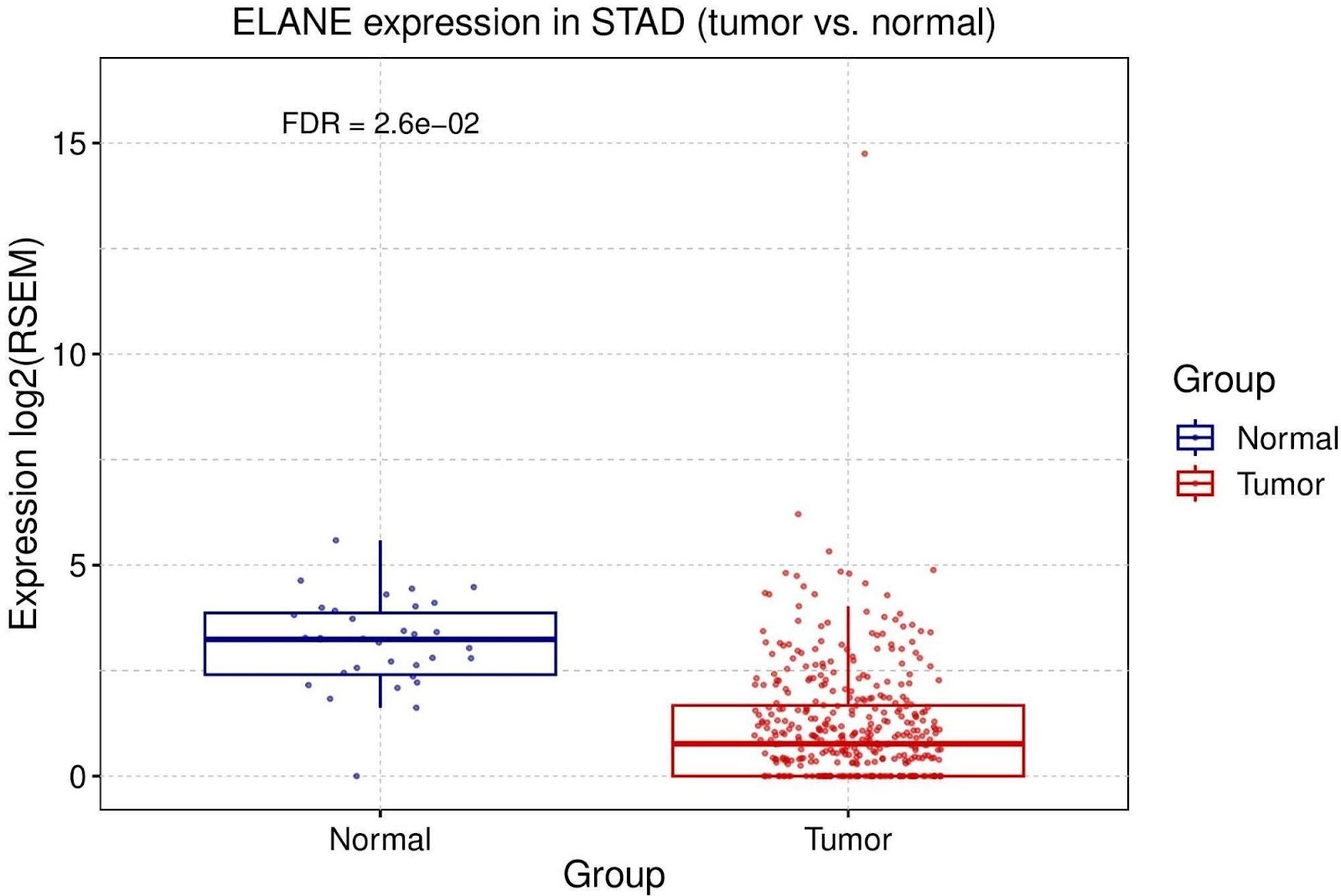
GC mRNA Expression & Clinical Survival Analysis of Pyroptosis Gene Set
In addition, comparative survival analysis was also conducted to provide a profile of the association of pyroptosis-related cell death genes and disease-free interval, disease-specific
survival, overall survival, and progression-free survival. Survival analysis showed that some pyroptosis genes have a relationship between expression and STAD survival. When overexpressed in STAD, ZEB2, NLRP1, GBP2, and ELANE appear to have a larger gastric
cancer tumor survival risk in comparison to other genes (Figure 3). ZEB2 appears to have the most statistically significant survival difference across multiple variations of survival as
Kaplan–Meier curves for disease-free interval, disease-specific survival, overall survival, and progress-free survival all demonstrate lower survival when ZEB2 displays higher expression levels (Figure 4). While there showed some significance across various genes such as STAD, NLRP1 and GBP2, their expression levels may not be critical for gastric cancer. These results indicated that the regulated expression of pyroptosis cell death genes might be involved in tumorigenesis and tumor development.
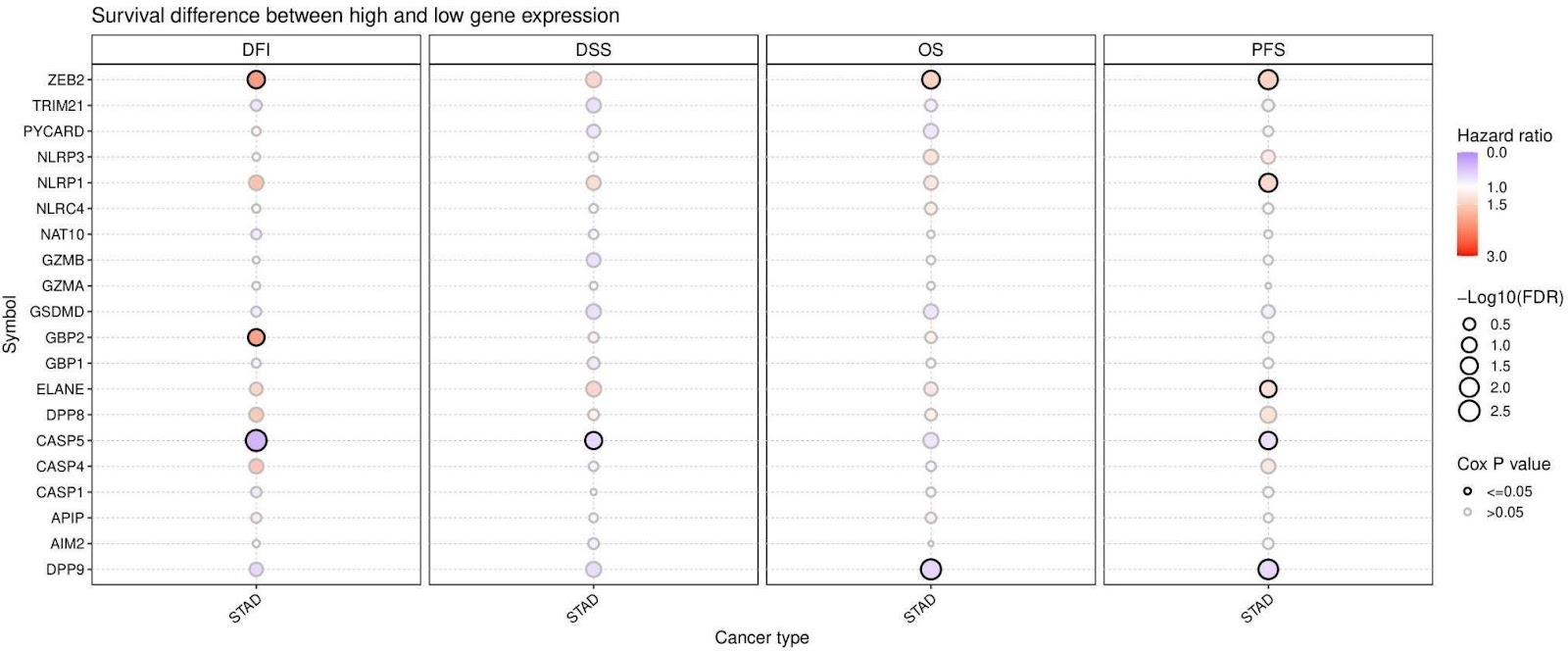
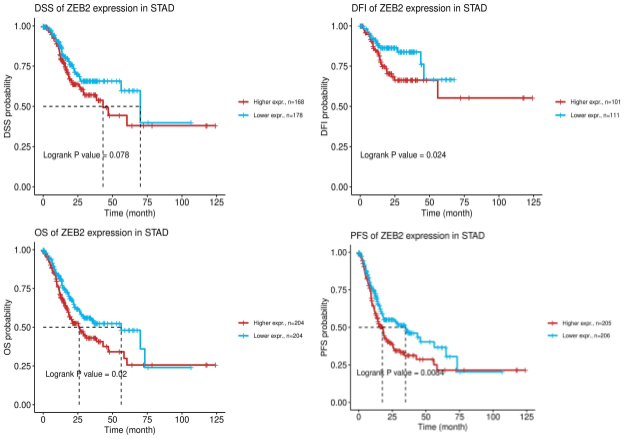
Discussion
In this study, the singular cancer analysis of the pyroptosis pathway associated with inflammatory response and bacterial infection showed its potential value as gastric cancer prognostic and diagnostic biomarkers. Expression analysis results regarding differential mRNA regulation of expression displayed that ELANE had the most significant differences, being one out of two genes that appeared to be down-regulated in GC. This is not in accordance with a prior study, where ELANE was found to be significantly up-regulated in GC and higher
expression of ELANE in GC cells was associated with pyroptosis. ELANE, which encodes neutrophil elastase (NET), is located on chromosome 19 and is one of six elastase genes found on humans. High NET plasma levels are known to be significantly related to pyroptosis as neutrophil elastases contained within NETs can significantly damage surrounding cells. During cases of infection, large amounts of ELANE are produced which could be involved in tumor
progression, poor patient survival, and reduced tumor prognosis on the basis of increased pyroptosis, relating to ELANE’s involvement in promoting inflammasome activation and
contributing to inflammatory responses37. However, touching back upon the dual-nature of the pyroptosis pathway in tumor development dependent on the surrounding tumor microenvironment and other biological factors, the researcher believes that it is possible for ELANE to be regulated in both directions that vary from different collected tumor samples, as inflammation. According to a study conducted by Du et. al (2021), moderate pyroptosis facilitates cells to maintain homeostasis, improve immune system activity, and effectively clear pathogens, all benefiting antitumor immunotherapy as such in the context of H.pylori-correlated GC38. However, excessive inflammation caused by pyroptosis is unfavorable to the host and may significantly aggravate tumor progression, which releases various inflammatory factors and forms an inflammatory immune microenvironment38. In the context of the dual role of the pyroptosis pathway in GC, there is not much existing literature about ELANE’s involvement.
However, high levels of pyroptosis are strongly correlated with high levels of neutrophil
elastases such as ELANE which additionally contributes to an overly inflammatory immune microenvironment and excessive inflammation surrounding cancer cells. ELANE could be a potential biomarker for GC. But as the differential mRNA expression and tumor survival varies across studies, future studies are strongly suggested to pursue further research on ELANE’s
expression regulation and the tendency of pyroptosis to aid in tumorigenesis rather than killing tumor cells.
Beyond ELANE down-regulation found in GC, ZEB2 has experienced down-regulation in the pyroptosis pathway of GC in multiple variants of survival with Hazard Ratio values greater than or equal to 1.44. According to our analysis, higher expression of ZEB2 correlates with higher risk of death and lower survival rates. From this, it seems likely that ZEB2
down-regulation in GC, decreasing mRNA expression, increases GC tumor survival in patients.
The down-regulation in the ZEB2 gene proves to be relevant as it is primarily a regulator of astrogliosis after ischemia or reperfusion injury, which enhances neuronal proliferation and
regeneration by reducing pyroptosis39. Thus, decreased expression or knockdown of ZEB2 in GC increases pyroptosis activity. In the context of gastric cancer, ZEB2 acts as a crucial transcription factor in EMT-driven chemoresistance, causing the breakdown of adhesion structures between tumor cells40. This enhances tumor invasiveness, contributes to chemoresistance, and results in treatment failure40. This indicates that in this study, increased down-regulation of ZEB2 in the pyroptosis pathways of GC indicates both higher pyroptosis and decreased EMT-mediated
chemoresistance, hinting that pyroptosis in GC may contribute to increased tumor progression and resistance to treatment. Thus, the ZEB2 gene in pyroptosis is a potential biomarker for GC
and has a potential use as a target in cancer therapies. This is a novel discovery and has not been reported in other studies. It does, however, support the tumorgenesis theory of pyroptosis brought about by this study’s analysis of ELANE, adding to prior studies identification of pyroptosis’ dual role in cancer progression. Future research is proposed to delve deeper into this relationship.
In the light of these two potential biomarkers highlighted within this study, it is worth addressing current well-established GC biomarkers as well. One of the most well-studied biomarkers of gastric cancer is HER2, which is not a part of the pyroptosis inflammatory
cell-death pathway. HER2 (Human Epidermal Growth Factor Receptor 2) is a member of the tyrosine kinase receptor family. When HER2 dimerizes, it triggers autophosphorylation of tyrosine residues in the receptor’s cytoplasmic domain, activating multiple signaling pathways that promote cell proliferation and tumor formation21. This receptor is widely expressed across various tissues, where it primarily supports uncontrolled cell growth and tumor development. HER2 amplification or overexpression occurs in approximately 10-30% of gastric cancers, making it a critical prognostic and predictive biomarker that informs targeted treatment
strategies21. Unlike HER2, the potential biomarkers ELANE and ZEB2 are not widely used as therapeutic targets and currently provide insights into tumor aggressiveness and potential immune interactions or metastatic risk. ELANE’s effects on GC through the creation of an inflammatory microenvironment still requires additional research since its expression may vary significantly based on the tumor microenvironment and other inflammatory processes, making it less specific compared to HER2. As for ZEB2, this study along with other academic literature have found its amplification consistently results in a higher risk of metastasis and pro-tumor growth, making it an ideal prognostic biomarker. Both ELANE and ZEB2 still require further inquiry, with possible applications in research and future implications in targeted interventions rather than standard care.
This study has a few potential limitations. For example, the Gene Set Cancer Analysis tool and TCGA-collected data do not take into account age or stage of cancer, which are clinical variables that may have affected the results of these analyses. Although this data has been
RSEM-normalized for more accurate comparison amongst differing samples, this factor is not
completely controlled for. Additionally, this study only analyzed tissue samples, which provide a static view of the cancer rather than capturing its dynamic progression. Another limitation is that the study focused solely on gastric cancer. Further research is needed to investigate bacterial infection-related cancers beyond gastric cancer to better understand pyroptosis-related gene sets in cancer development. Despite these limitations, the study offers valuable insights into potential diagnostic and prognostic biomarkers and targeted therapies in clinical settings. These findings promote additional research into the ELANE and ZEB2 as gastric cancer biomarkers, which in the clinical setting, enhances early detection, improves screening, therapeutic efficacy, and prediction of recurrence risks and metastasis, further enabling better treatment and recovery for GC patients. Further studies similarly exploring the field of using genetic profiles and cancer will pave a stronger path for a clinical future of precision medicine, where biomarkers can identify
specific molecular targets within tumors, leading to more effective treatments. For instance, targeting a gene involved in gastric cancer proliferation or survival can help inhibit cancer growth with fewer side effects compared to conventional therapies.The findings also present a comprehensive overview of the pyroptosis pathway gene set, which can serve as a reference for future cancer research.
Conclusion
This study thoroughly explored the genomic cancer profiles of the pyroptosis gene set by analyzing mRNA expression and survival data across over 450 samples and 32 paired samples of stomach adenocarcinoma. The results highlighted ELANE and ZEB2 as genes with reduced
expression in pyroptosis, suggesting that gastric cancer might be influenced by this process. The analysis of pyroptosis-related genes in gastric cancer could offer new insights for developing
clinical therapies that target pyroptosis. However, more extensive pan-cancer research is necessary to investigate additional genes and evaluate the potential of pyroptosis as a biomarker across various cancer types beyond gastric cancer.
References
- Siegel, R. L., Giaquinto, A. N., & Jemal, A. Cancer statistics, 2024. CA: A Cancer Journal for Clinicians. 74, 12–49 (2024). [↩]
- Collins, S. 2024–First year the US expects more than 2M new cases of cancer. American Cancer Society. (2024). https://www.cancer.org/research/acs-research-news/facts-and-figures-2024.html [↩]
- Matsuoka, T., Yashiro, M. Biomarkers of gastric cancer: Current topics and future perspective. World Journal of Gastroenterology. 24, 2818–2832 (2018). [↩]
- Ho, S.W.T., Tan P. Dissection of gastric cancer heterogeneity for precision oncology. Cancer Science. 110, 3405-3414 (2019). [↩]
- Puntmann, V. O. How-to guide on biomarkers: biomarker definitions, validation and applications with examples from cardiovascular disease. Postgraduate Medical Journal. 85, 538–545 (2009). [↩]
- Peng, F., Liao, M., Qin, R., Zhu, S., Peng, C., Fu, L., Chen, Y., Han, B. Regulated cell death (RCD) in cancer: key pathways and targeted therapies. Signal Transduct and Target Therapy 7, 286 (2022). [↩]
- Jia, Y., Wang, X., Deng, Y., Li, S., Xu, X., Qin, Y., & Peng, L. Pyroptosis provides new strategies for the treatment of cancer. Journal of Cancer. 14, 140–151 (2023). [↩] [↩] [↩] [↩] [↩]
- Li, L., Dickinson, M.S., Coers, J., Miao, E.A. Pyroptosis in defense against intracellular bacteria. Seminars in Immunology. 69, (2023). [↩]
- Wei, X., Xie, F., Zhou, X., Wu, Y., Yan, H., Liu, T., Huang, J., Wang, F., Zhou, F., Zhang, L. Role of pyroptosis in inflammation and cancer. Cellular & Molecular Immunology. 19, 971-992 (2022). [↩] [↩]
- Burdette, B., Esparza, A., Zhu, H., Wang, S. Gasdermin D in pyroptosis. Acta Pharmaceutica Sinica B. 11, 2768-2782 (2021). [↩]
- Grivennikov, S., Greten, F., Karin, M. Immunity, inflammation, and cancer. Cell. 140, 883-899 (2010). [↩] [↩] [↩]
- Yin, Q., Song, S., Bian, Y., Wang, Y., Deng, A., Lv, J., Wang, Y. Unlocking the potential of pyroptosis in tumor immunotherapy: a new horizon in cancer treatment. Front Immunology. 15, (2024). [↩]
- Moss, S. The Clinical Evidence Linking Helicobacter pylori to Gastric Cancer. Cell Mol Gastroenterol Hepatol. 3, 183-191 (2016).; Bacteria that Can Lead to Cancer. American Cancer Society. (2024). https://www.cancer.org/cancer/risk-prevention/infections/infections-that-can-lead-to-canc er/bacteria.html [↩]
- Hong, S., Lee, W. J., Kim, Y. D., Kim, H., Jeon, Y., Lim, B., Cho, D., Heo, W. D., Yang, D., Kim, C., Yang, H., Yang, J. K., & Jung, Y. APIP, an ERBB3-binding partner, stimulates erbB2-3 heterodimer formation to promote tumorigenesis. Oncotarget. 7, 21601–21617 (2016). [↩]
- Wei, S., Feng, M., & Zhang, S. Molecular characteristics of cell pyroptosis and its inhibitors: A review of activation, regulation, and inhibitors. International Journal of Molecular Sciences. 23, 16115 (2022). [↩]
- Lu, L., Zhang, Y., Tan, X., Merkher, Y., Leonov, S., Zhu, L., Deng, Y., Zhang, H., Zhu, D., Tan, Y., Fu, Y., Liu, T., & Chen, Y. Emerging mechanisms of pyroptosis and its therapeutic strategy in cancer. Cell Death Discovery. 8, 338 (2022). [↩] [↩] [↩]
- Hollingsworth, L.R., Sharif, H., Griswold, A.R., Fontana, P., Mintseris, J., Dagbay, K.B., Paulo, J.A., Gygi, S.P., Bachovchin, D.A., & Wu, H. DPP9 sequesters the C terminus of NLRP1 to repress inflammasome activation. Nature. 592, 778–783 (2021). [↩]
- Wang, X., Zhao, Y., Wang, D., Liu, C., Qi, Z., Tang, H., Liu, Y., Zhang, S., Cui, Y., Li, Y., Liu, R., & Shen, Y. ALK-JNK signaling promotes NLRP3 inflammasome activation and pyroptosis via NEK7 during Streptococcus pneumoniae infection. Molecular Immunology. 157, 78–90 (2023). [↩]
- Damiano, J.S., Oliveira, V., Welsh, K., & Reed, J.C. Heterotypic interactions among NACHT domains: implications for regulation of innate immune responses. Biochemical Journal. 381, 213–219 (2004). [↩]
- Fernandes-Alnemri, T., Yu, J., Datta, P., Wu, J., & Alnemri, E.S. AIM2 activates the inflammasome and cell death in response to cytoplasmic DNA. Nature. 458, 509–513. (2009). [↩]
- Iqbal N, Iqbal N. Human Epidermal Growth Factor Receptor 2 (HER2) in Cancers: Overexpression and Therapeutic Implications. Mol Biol Int. 2014, (2014). [↩] [↩] [↩]
- McConnell, B.B., Vertino, P.P. Activation of a caspase-9-mediated apoptotic pathway by subcellular redistribution of the novel caspase recruitment domain protein TMS1. Cancer Research. 60, 6243-6247 (2000). [↩]
- Ghosh, A., Praefcke, G.J.K., Renault, L., Wittinghofer, A., & Herrmann, C. How guanylate-binding proteins achieve assembly-stimulated processive cleavage of GTP to GMP. Nature. 440, 101–104 (2006). [↩]
- Braun, E., Hotter, D., Koepke, L., Zech, F., Grob, R., Sparrer, K., Muller, J., Pfaller, C., Huesinger, E., Sutter, K., Dittmer, U., Winkler, M., Simmons, G., Jakobsen, M., Conzelmann, K., Pohlman, S., Munch, J., Fackler, O., Kirchhoff, F., Sauter, D. Guanylate-Binding Proteins 2 and 5 Exert Broad Antiviral Activity by Inhibiting Furin-Mediated Processing of Viral Envelope Proteins. Cell Reports. 27, 2092-2104 (2019). [↩]
- Kang, W., & Yang, J. K. Crystallization and preliminary X-ray crystallographic analysis of human Apaf-1-interacting protein. Acta Crystallographica Section F Structural Biology and Crystallization Communications. 68, 1518–1520 (2012). [↩]
- Zhou, Z., He, H., Wang, K., Shi, X., Wang, Y., Su, Y., Wang, Y., Li, D., Liu, W., Zhang, Y., Shen, L., Han, W., Shen, L., Ding, J., Shao, F. Granzyme A from cytotoxic lymphocytes cleaves GSDMB to trigger pyroptosis in target cells. Science (New York, N.Y.) 368, eaaz7548 (2020). [↩]
- Poe, M., Blake, J.T., Boulton, D.A., Gammon, M., Sigal, N.H., Wu, J.K., Zweerink, H.J. Human cytotoxic lymphocyte granzyme B. Its purification from granules and the characterization of substrate and inhibitor specificity. The Journal of Biological Chemistry. 266, 98-103 (1991). [↩]
- Zhang, H., Chen, Z., Zhou, J., Gu, J., Wu, H., Jiang, Y., Gao, S., Liao, Y., Shen, R., Miao, C., & Chen, W. NAT10 regulates neutrophil pyroptosis in sepsis via acetylating ULK1 RNA and activating STING pathway. Communications Biology. 5, 916 (2022). [↩]
- Xue, B., Li, H., Guo, M., Wang, J., Xu, Y., Zou, X., Deng, R., Li, G., & Zhu, H. TRIM21 Promotes Innate Immune Response to RNA Viral Infection through Lys27-Linked Polyubiquitination of MAVS. Journal of Virology. 92, e00321-18 (2018). [↩]
- Johnson, D.C., Taabazuing, C.Y., Okondo, M.C., Chui, A.J., Rao, S.D., Brown, F.C., Reed, C., Peguero, E., De Stanchina, E., Kentsis, A., & Bachovchin, D.A. DPP8/DPP9 inhibitor-induced pyroptosis for treatment of acute myeloid leukemia. Nature Medicine. 24, 1151–1156 (2018). [↩] [↩]
- Zhao, S., Tan, Y., Qin, J., Xu, H., Liu, L., Wan, H., Zhang, C., Fan, W., & Qu, S. MicroRNA-223–3p promotes pyroptosis of cardiomyocyte and release of inflammasome factors via downregulating the expression level of SPI1 (PU.1). Toxicology. 476, 153252 (2022). [↩]
- Qi, S., Song, Y., Peng, Y., Wang, H., Long, H., Yu, X., Li, Z., Fang, L., Wu, A., Luo, W., Zhen, Y., Zhou, Y., Chen, Y., Mai, C., Liu, Z., & Fang, W. ZEB2 mediates multiple pathways regulating cell proliferation, migration, invasion, and apoptosis in glioma. PLOS ONE. 7, e38842 (2012). [↩]
- Liu, C., Hu, F., Xie, G., Miao, Y., Li, X., Zeng, Y., Guo, A. GSCA: an integrated platform for gene set cancer analysis at genomic, pharmacogenomic and immunogenomic levels, Briefings in Bioinformatics. 24, (2023). [↩]
- Liu, C., Hu, F., Xie, G., Miao, Y., Li, X., Zeng, Y., Guo, A. GSCALite: a web server for gene set cancer analysis. Bioinformatics. 34, (2018). [↩]
- Liu, H. Pan-cancer profiles of the cuproptosis gene set. American Journal of Cancer Research. 12, 4074-4081 (2022). [↩]
- Liu, H., & Tang, T. Pan-cancer genetic analysis of cuproptosis and copper metabolism-related gene set. Frontiers in Oncology. 12, 952290 (2022 [↩]
- Cui, M., Wang, X., Qiao, H., Wu, S., & Shang, B. ELANE is a Promising Prognostic Biomarker that mediates Pyroptosis in Gastric Cancer. Heliyon. 10, e34360 (2024). [↩]
- Du, T., Gao, J., Li, P., Wang, Y., Qi, Q., Liu, X., Li, J., Wang, C., Du, L. Pyroptosis, metabolism, and tumor immune microenvironment. Clinical and Translational Medicine. 11, 492 (2021). [↩] [↩]
- Vana, F., Szabo, Z., Masarik, M. Kratochvilova, M. The interplay of transition metals in ferroptosis and pyroptosis. Cell Div 19, (2024). [↩]
- Yuan, L., Xu, ZY., Ruan, S.M., Mo, S., Qin, J.J., Cheng, X.D. Long non-coding RNAs towards precision medicine in gastric cancer: early diagnosis, treatment, and drug resistance. Molecular Cancer 19, (2020). [↩] [↩]

 and Family-Integrated Care (FIC): Global Trends and Local Provider Awareness in Fresno County, California")

{kind=link}