
Abstract
Plastic pollution is a major environmental challenge that impacts ecosystems and human health. Traditional chemical based recycling methods are ineffective, leaving a large amount of plastic either piled up in landfills or floating in the ocean for centuries. Enzymatic degradation of plastics offers a sustainable alternative to recycle plastics. PETase, the enzyme capable of degrading polyethylene terephthalate (PET), can become a new biotechnological workhorse. This report explores the production of PETase using recombinant technologies. We investigated multiple heterologous systems for protein expression, including Rosetta and BL21 E. coli strains and Sf9 insect cells. Protein expression was scaled using both shake flasks and a 5-liter bioreactor. Rosetta strains demonstrated moderate yields at around 17mg/L or 1.5mg/g of biomass. Yield per liter was increased to 150mg/L when using Sf9 cells. Notably, 5-liter bioreactor cultivation using E. coli maintained a similar 1.5mg/g of biomass yield but a significant increase of total protein yield per liter of culture to approximately 430mg/L. These results highlight feasibility of further scalable industrial applications, and support its potential integration into a commercial recycling system for enzymatic plastic biodegradation. PETase offers a more effective and efficient way for dealing with the plastic crisis.
Keywords: Plastic Pollution, Protein Production, Enzymes, PETase, Biotechnology/Synthetic Biology, Heterologous Systems, Biodegradation
Introduction
Plastic pollution plagues the world as one of the most urgent problems that humanity has to deal with. Compounds that are often found in our plastic bottles can leach into the water, causing serious health issues like hormone changes, insulin resistance, and cancer1. These microplastics, or plastic particles that come from the natural erosion of larger plastic waste, are everywhere in the soil in which we grow crops and subsequently consume. The 11 million tons of plastics that accumulate in oceans every year also harm marine life and the current recycling solutions are ineffective2. For instance, the standard single-stream recycling process has existed since the 1990s, and it is what most of the world uses today. This 4-step process, including collecting, transporting, sorting, and recycling the plastics, has not been effective in the past and will not be in the future3. 80% of recyclable materials end up in landfills because of clumsy sorting issues or the tolerance of the recycling plant for mixed materials4. When plastics end up in landfills, it takes centuries for it to degrade. For these reasons, new methods to deal with the consequences of plastic pollution are a priority.
Enzymatic degradation of plastic can overcome the shortcomings of the current chemical and mechanical recycling methods5,6,7,8. Some of the disadvantages of the mechanical recycling system generally include the over reliance on human sorting and limitations of the recycling plants3. Chemical recycling solutions include gasification and incineration. Incineration, though quick and effective, produces over 0.7-1.7 tonnes of carbon dioxide for every tonne of plastic9. Gasification does not have the disadvantages of the fumes, but rather have really high initial costs and high specificity of the moisture, inert content, and calorific value10.
Biotech researchers have discovered and further developed enzymes called PETase enzymes that can biologically degrade plastics made of PET (Polyethylene Terephthalate), organically6,7,8. In Table 1, it is shown that enzymatic degradation of plastics has a lower carbon footprint than conventional methods. In addition, enzymes reduce the reliance on humans, effectively decreasing cost and labor complexities. Enzymes can be engineered to recognize and depolymerize specific polymers (for example, PET-targeting hydrolases), so mixed waste streams that are only coarsely separated by type can still be processed effectively, reducing how much fine, item-by-item sorting humans must do5. There are also fewer requirements for the specific temperature and pressure in which the enzyme can operate as its reaction conditions are mild11. The associated costs for PETase enzymes are research and development and scale up costs, which can be greatly reduced in the near future as technology matures. PETase enzymes can greatly increase the efficiency of existing recycling systems and reduce costs when fully developed.
This study seeks to design and evaluate scalable recombinant expression platforms for PETase, with the overarching goal of enabling its deployment as an industrial biocatalyst for enzymatic PET recycling. While numerous PETase variants have been engineered with enhanced catalytic activity and thermostability, suboptimal expression levels and solubility in conventional heterologous hosts remain a critical bottleneck for large-scale implementation12. To address this constraint, the work systematically benchmarks PETase production across distinct expression modalities—E. coli shake flasks, Sf9 insect cell cultures, and a 5 L E. coli bioreactor—quantifying performance in terms of biomass-specific yield (mg/g) and process suitability for cost-effective intensification. Nevertheless, because PETase yields in E. coli are moderate, we explored Sf9 cells, a widely used insect cell line in biotechnology derived from Spodoptera frugiperda. Sf9 cells are eukaryotic, which allows them to fold complex proteins correctly13 and perform post-translational modifications (PTMs), such as glycosylation and disulfide bond formation, that are critical for protein stability, solubility, and enzymatic activity14,15,16. This makes them particularly useful for expressing proteins like PETase that may require such modifications to achieve optimal function13. By demonstrating that optimized prokaryotic fed-batch bioreactor cultivation can markedly increase PETase productivity, this study delineates a viable bioprocessing route toward industrially relevant PETase manufacturing.
Methods
Bacterial Transformation Using Expression Vector
We used three constructs, all cloned in pET28 derivative backbones. The constructs contain a green fluorescent protein (pET-GFP), a nanobody (pET-Nano) and a version of PETase (pET-PETase) which are under the control of the T7 polymerase promoter. The PETase construct contains the full-length enzyme sequence fused to a C-terminal hexahistidine (His₆) tag via a GGS linker. The gene was codon-optimized for expression in E. coli, synthesized by GenScript, and subcloned into a pET28-derivative expression vector, which was used to transform the E. coli strains described in this study. First, chemically competent E. coli cells of the BL21 and Rosetta strains are obtained from -80°C freezer and thawed on ice. The thawed 25 ul of competent cells are mixed with 100 ng of plasmid DNA of each of the constructs (pET-GFP and pET-PETase) and incubated on ice for 30 minutes. After incubation on ice, a heat shock at 42°C for 45 seconds was applied. The mixture of cells and plasmid DNA was immediately returned to ice to facilitate DNA uptake for 5 minutes. After this second incubation on ice, 200-250 ul of SOC medium was added for recovery and the cells were immediately incubated at 37°C in a shaker at 200 rpm for 1h. Lastly, the cells are plated onto LB agar plates containing kanamycin in 50ug/ml (Kan) concentration for selection.
Small scale and Large Scale Protein Expression In Shake Flasks (E. coli)
Small scale expressions were performed at a volume of 3mL. For these expressions, 1mL of precultures in LB supplemented with Kanamycin (50ug/mL) were incubated overnight at 37°C and 200 rpm. Next day, 100uL of the preculture was used to inoculate 3mL of either TB media or Autoinduction. TB media was incubated at 37°C with shaking (200 rpm) until the Optical Density (OD) reached 0.6-0.8, induced with IPTG (0.5mM), temperature reduced to 30°C, and expressed for 16-18h. For autoinduction, we followed a similar protocol with the same volumes but without expression with IPTG and only growing at 25°C (without temperature switch) for the duration of the expression (16-18h). For the large scale expressions, 200mL of preculture using LB supplemented with Kanamycin (50ug/mL) was incubated overnight at 37°C and 200 rpm. After approximately 16h of incubation the preculture was visually inspected for signs of contamination prior to inoculation. For the main culture, cells were grown in 2.5 L shake flasks. Each 2.5 L flask was filled with 1 L of TB media, supplemented with Kan (50ug/mL), and a 1/100 volume of pre-culture was added. After sealing the flasks with breathable tape, they were incubated at 37°C with shaking (200 rpm). Optical density readings were taken at the time of incubation and after 1 hour with subsequent readings every 30 minutes. When the OD reached the target range of 0.6-0.8, the IPTG was added to initiate protein expression (0.5mM), and the incubation temperature set to 30°C. Culture was induced for 16h.
Insect Cell Expression
Fresh bacmid DNA, containing the recombinant baculoviral genome encoding the PETase gene, was transfected into log-phase Sf9 cells to generate the initial baculovirus stock (P0). Sf9 cells were plated at 0.5 × 10⁶ cells/mL and transfected using CellFectin II under multiple transfection conditions, including appropriate controls. After 5 hours, the transfection mixture was replaced with supplemented medium, and cells were incubated at 27 °C. Virus-containing supernatant was harvested 5 days post-transfection to obtain the P0 baculovirus stock.
To amplify the virus, log-phase Sf9 cells were diluted to 0.5 × 10⁶ cells/mL and plated into 6-well dishes. Ten microliters of P0 virus were added per well, and cells were incubated at 27 °C. After 5 days, the supernatant was collected to generate the P1 baculovirus working stock.
A small-scale expression test was then performed in HiFive cells, which are used for high-level recombinant protein expression and secretion in the baculovirus system. These cells also provide eukaryotic post-translational modifications, such as glycosylation, which can be important for proper folding and activity of secreted proteins. HiFive cells were diluted to 0.5 × 10⁶ cells/mL and plated into 6-well dishes, followed by infection with P1 virus at dilutions ranging from 1:40 to 1:400, along with an uninfected control. Cells were incubated for 48 hours, after which they were collected.
Sf9 and HiFive insect cell lines were cultured at 27 °C with shaking at 140 rpm, with vessel caps loosened to allow aeration. Sf9 cells were maintained at densities of 1.0–2.0 × 10⁶ cells/mL and subcultured every other day in insect medium supplemented with 10% FBS, while HiFive cells were maintained at 0.8–2.5 × 10⁶ cells/mL and subcultured daily in Express Five SFM. Frozen cell stocks were thawed in a 37 °C water bath prior to use. Cell density and viability were monitored daily, maintaining cultures at a final volume of 50 mL and approximately 1.0 × 10⁶ cells/mL.
Large Scale Protein Expression In E. coli Using Bioreactors
Escherichia coli BL21 (DE3) transformed with the pET28 vector carrying the PETase gene was utilized for this experiment. Long-term preserved glycerol stocks (30% v/v) were used to prepare the seed cultures (250mL) which were cultivated in rich TB media and grown at 37°C until the stationary phase was reached. Prior to bioreactor inoculation, cells from the seed culture were harvested by centrifugation, and resuspended in a defined mineral salt medium to prevent metabolites carryover and grown for a couple of hours at 37°C. Cultivations were performed in a 5 L stirred-tank bioreactor. The initial batch phase utilized a defined complex medium containing glucose and salts, adjusted to an initial pH of 7.2. During the growth phase, the temperature was maintained at 37°C. The dissolved oxygen (DO) concentration was maintained at 30% of air saturation via a cascade control of agitation (up to 1200 rpm) and aeration. The pH was actively controlled through the automated addition of 2 M NaOH. To achieve high cell-density, a fed-batch strategy was implemented using a glucose-rich feeding ramp initiated upon the exhaustion of initial batch nutrients (indicated by a DO spike). Biomass accumulation was monitored via optical density measurements at 600 nm (OD600). Recombinant PETase expression was induced at an 𝑂𝐷600 of approximately 19 by the addition of 0.5 mM IPTG. Optical density measurements used to monitor biomass accumulation during cultivation, serving as an indirect proxy for cell density and growth. OD values were used to determine appropriate timing for induction and sampling during bioreactor operation. Simultaneously, the culture temperature was shifted to 30°C to facilitate proper protein folding and enhance solubility. Growth kinetics were tracked through frequent 𝑂𝐷600 sampling (every 30 min post-inoculation; every 2–3 h during the high-density phase) until a final 𝑂𝐷600 of 120 was achieved. Protein expression was characterized using Sodium SDS-PAGE followed by Coomassie Brilliant Blue staining. Target protein concentration was determined via densitometric analysis against known protein standards for PETase quantification.
Protein Purification Using Imac
We have used an established method of IMAC purification with minor alterations, which is described in this section17. Cell pellets were resuspended in a lysis buffer and disrupted by probe sonication using multiple short bursts with cooling periods; this minimizes heating and improves protein release for preparing cleared lysates for downstream affinity purification. An IMAC column filled with nickel-nitrilotriacetic acid (Ni-NTA) or cobalt-based resin was loaded with the cleared lysate after it had been pre-equilibrated with binding buffer that contained 10 mM imidazole, 300 mM NaCl, and 50 mM sodium phosphate (pH 7.5). To guarantee effective binding of the His-tagged protein to the immobilized metal ions on the resin, the lysate was added at a flow rate of 1 mL/min. The column volumes of the wash buffer comprising 50 mM sodium phosphate (pH 7.5), 300 mM NaCl, and 20–30 mM imidazole were used to wash the column after loading in order to get rid of proteins that were not precisely bound.
A step gradient or single elution buffer of 250 mM imidazole, 300 mM NaCl, and 50 mM sodium phosphate (pH 7.5) was used to elute the associated His-tagged protein. Elution was carried out. Elution was conducted at a controlled flow rate of 1 mL/min, and fractions of 1 mL were collected. The fractions were analyzed by SDS-PAGE to confirm the presence of the target protein.
Results
Transformation, And Small Scout To Test Expression In E. coli
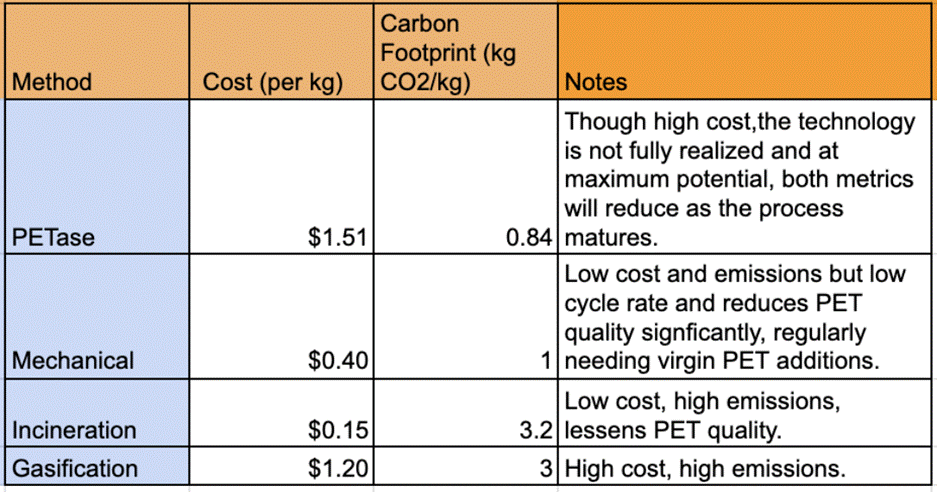
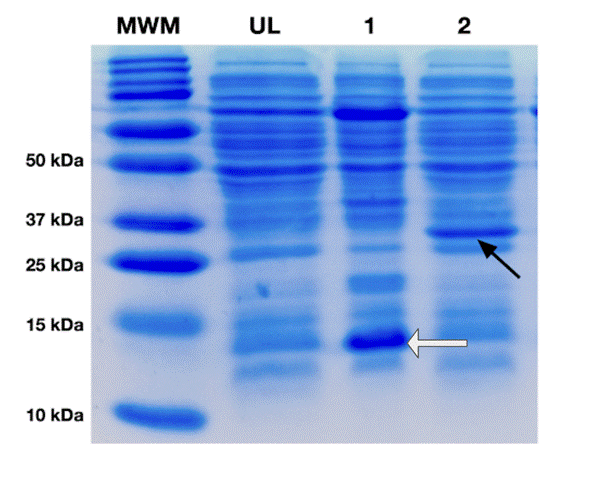
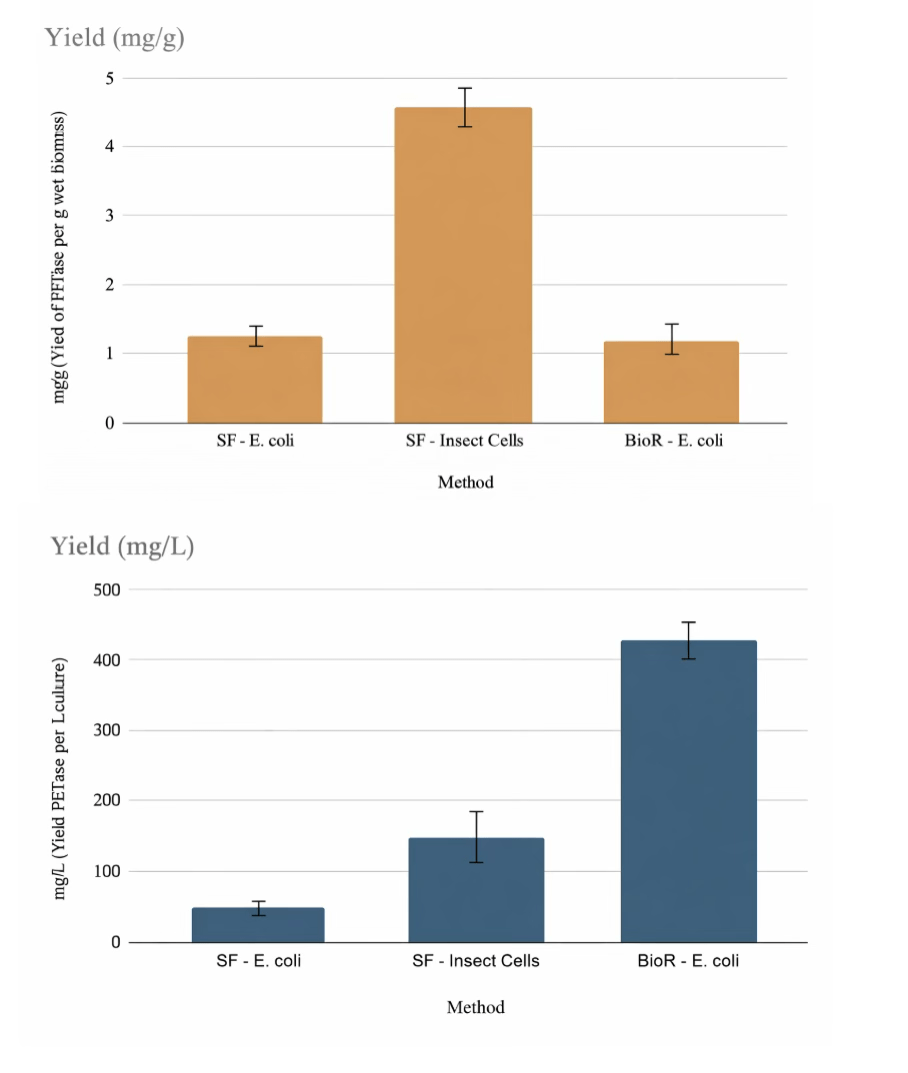
In order to test the expression of PETase, we performed the transformations of BL21 and Rosetta strains with a plasmid (derivative of pET28) containing a full-length version of PETase. As control for expression, we used GFP cloned in the same pET28 derivative vector and transformed into BL21 cells. After a successful transformation, evidenced by the growth of approximately 200 colonies on selective LB agar plates, we proceeded with a small expression scout (3mL) to test growth in two conditions, TB media induced with IPTG (1mM) at 30°C for 16h, and Autoinduction at 25°C for 16-18h. Initially both of the controls were only expressed in one condition (TB, induced with IPTG at 30°C for 16h)(figure 1) as TB was previously determined to work for both constructs (data not shown). We ran the cytosolic extract for both control expressions in an SDS-PAGE (Figure 1) the nanobody and GFP (lines one and two in Figure 1, respectively), expressed as expected, as demonstrated by the difference in the increase in the intensity of the bands corresponding molecular their respective molecular weights (around 14 KDa for the nanobody, and 27 kDa for GFP) when compared to the preinduced E. coli cytosol (Line UL in Figure 1, which represent the cytosolic extract before induction with IPTG). After expressing the PETase constructs in both conditions mentioned above, and even though the protein expressed similarly in all conditions as seen in Figure 2, the scout results showed a slight improvement of expression using the Rosetta strain in Autoinduction (line 4 in Figure 2), confirmed by SDS-PAGE densitometry analysis using the ImageJ software, although all of the expressions showed a prominent band at the target protein’s predicted molecular weight (27kDa) after cell lysis and protein extraction (Figure 2). A semi-quantification by densitometry, analyzing the SDS-PAGE image with the program Image J, shows that our conditions produce approximately 15mg/L of PETase or approximately 1.5mg of enzyme per gram of Biomass (calculated by correlating the mass of the His-tag purified protein with the corresponding wet biomass weight), which is consistent with previous reports18 and is considered a moderate yield. Nevertheless, as the differences do not seem to be significant, and in order to have a better control of the expression and fit for the bioreactor protocol, we decided to move forward with the TB condition induced by IPTG using the Rosetta strain (30°C).
Scale Up In Shake Flasks And Pilot Protein Purification
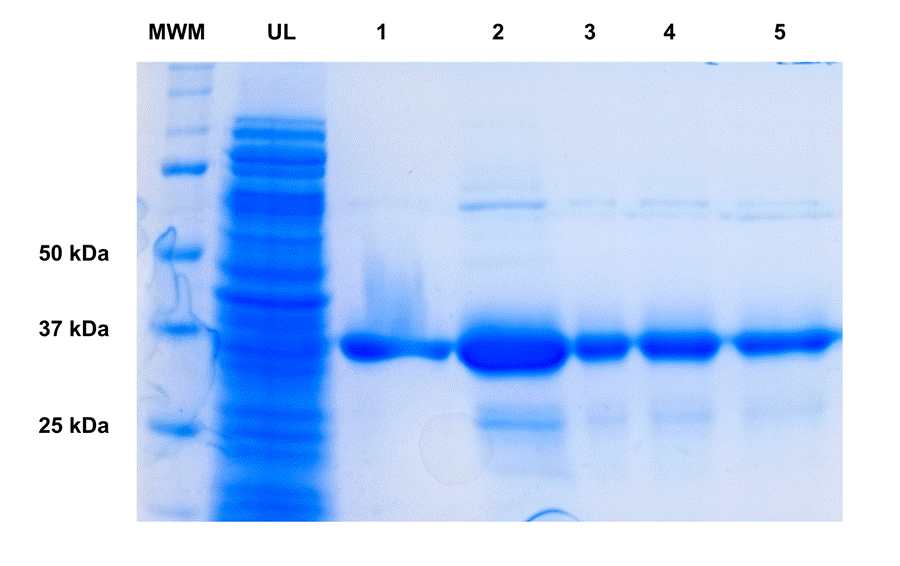
Using the conditions determined by the scout (Rosetta strain and TB media at 30°C), we scaled up the expression of PETase to 1L in shake flasks. The expression in larger volumes showed similar results compared to the small scout with a semi-quantification by densitometry yielding around 17mg/L (or approximately 1.5mg/g of biomass) under these optimized conditions. Given the good results of both the scout and large scale expression (1L) of the PETase expression using the Rosetta strain and Autoinduction, we moved forward with 1 step purification using Immobilized Metal Affinity Chromatography (IMAC). The biomass output, which amounted to roughly 10–12 grams of wet cell weight, was processed using an emulsifier and the lysate loaded onto the IMAC. The target protein was successfully separated from other impurities by eluting with a step gradient of imidazole in buffer Tris as described in the methods section. The eluted fractions analyzed by SDS-PAGE revealed a noticeable band that matches the PETase molecular weight (Figure 3). The purification yielded a total of 13mg of protein from 1L culture with a final concentration of 1.3mg/mL and a purity around 90% as calculated by densitometry.
Expression Using Insect Cells And Large Scale Expression Of E. coli Using Bioreactors
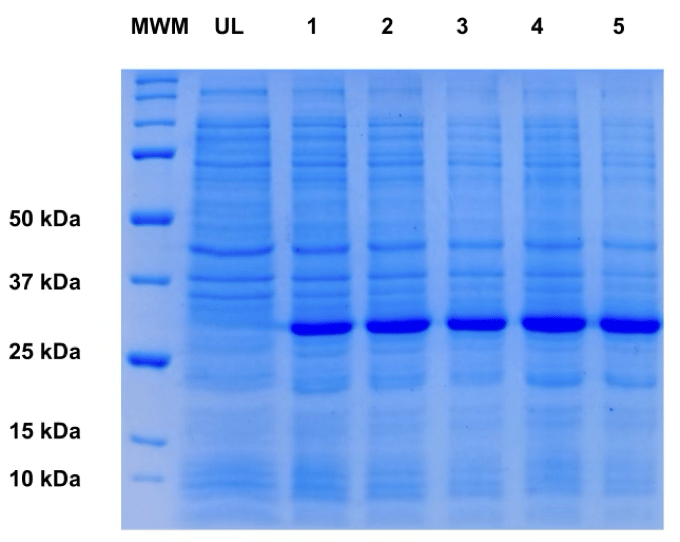
Given the moderate expression observed in E. coli, we decided to test expression in Sf9 cells that were cultivated in shake flasks after being infected with a recombinant baculovirus for expression in insect cells19. Shown in figure 4, the yields for the secreted PETase obtained from Sf9 cell cultures was 150mg/L of culture, a significant improvement over the yield from the E. coli system. This result demonstrates the superiority of eukaryotic cells to express PETase.
Nevertheless, the complexity and cost of eukaryotic cell cultures made us pivot to increasing the yields or total production using E. coli through the application of Bioreactors. During the fermentation process, the bacterial biomass and the enzymatic yields were calculated at the end of the run. The yields observed within the 5-liter bioreactor were similar to shake flasks in terms of mass of protein over biomass (around 1.5mg/g). However, the biomass produced by the bioreactor was 10 times higher, around 80g/L of culture and with a final OD of 85, compared to shake flasks (OD of 9 and around 10-12g of biomass). The superior biomass production of the Bioreactor pushed the enzyme production yields per liter to around 450mg/L.
Discussion
Degradation and recycling of PET plastics by enzymatic means is a promising solution to alleviate the harms of plastic pollution. The application of such green technologies can be not only cost-effective but less labor intensive than existing solutions and more environmentally friendly and sustainable. However, some problems that exist include the development of techniques that can degrade different types of plastics (and mixture of plastics) and limitations of scale-up efforts due to the relatively poor expression of PETase enzymes in heterologous systems compared to other industrial enzymes20,21.
In this study, we addressed the latter challenge by demonstrating a scalable production system for PETase. We were able to produce up to 450mg/L of PETase using a strain of E. coli and a 5L Bioreactor. Even though these yields are still moderate in comparison to other industrial enzymes (1-10g/L)20, it shows that PETase can be successfully expressed in prokaryotic cells. While expression in Sf9 cells cultivated in shake flasks produced a comparable yield per liter, this platform, although advantageous in terms of downstream processing due to secretion of the enzyme, is associated with higher cultivation costs. Under the conditions used in this study, the estimated culture cost was approximately $63 per liter for E. coli and $110 per liter for Sf9 insect cells, reflecting the greater media and maintenance requirements of eukaryotic cell culture systems. Consequently, despite the benefits of secretion, insect cell expression remains less cost-effective than prokaryotic production for scalable PETase manufacturing
To place our results in context, it is useful to compare PETase production levels reported in similar heterologous expression systems. Recent work in E. coli has shown that using strains engineered to promote disulfide bond formation can yield approximately 20mg of purified PETase per liter of shake-flask culture, while moving to controlled bioreactor cultivation increased volumetric yield to around 10mg/L for IsPETase under optimized conditions22. Other variants of PETase, such as FAST-PETase and Hot-PETase, have also been produced in E. coli with purified yields on the order of 65–77mg/L12. In contrast, our E. coli shake-flask yields (~17mg/L , ~1.5mg/g biomass) are broadly consistent with the lower end of reported titers, and our 5 L bioreactor yields (~430–450mg /L ) represent a marked increase compared with many shake-flask studies, underscoring the benefit of high-cell-density bioreactor cultivation. Notably, while there are fewer reports of PETase production in insect cells, eukaryotic systems have been shown to improve expression and secretion for other recombinant proteins, and our ~150mg/L yield in HiFive cultures demonstrates that insect expression can outperform conventional E. coli shake-flask systems for PETase. Together, these comparisons highlight both the consistency of moderate yields in basic E. coli systems and the advantage of bioreactor and alternative host strategies for increasing total PETase output, while also pointing to a gap in standardized bioreactor data for direct comparison that our work helps to address.
Our findings suggest that the expression system and cultivation parameters on Bioreactors can be further optimized for PETase production, making this approach more promising for scale-up and industrial applications. Our production in Bioreactors yields 10% (g/L) of Biomass, with an OD of around 85 which is approximately 80g/L of culture. Further optimization of the production parameters, including the media and use of exponential feeding, could potentially yield even higher biomass production. Although the process to increase the biomass can still encounter several bottlenecks such as metabolic burden, expression toxicity, or oxygen transfer limits, it is reasonable to think that up to 20% to 25% of wet cell biomass per liter of culture can be achieved, which could potentially duplicate the enzyme yields per liter23,24,25. In addition, other prokaryotic systems such as Bacillus subtilis can be used as heterologous systems for the production of PETase and which combine the advantages of being cost-effective and having an efficient secreting system26.
Given the promising results presented in this study, there is a clear imperative for continued research and industrial investment in scalable PETase production systems. In parallel, interdisciplinary efforts integrating biochemical engineering, synthetic biology, and industrial-scale process design should be mobilized to bridge the current gap between laboratory-scale production and full-scale establishment. The urgency of the plastic pollution crisis demands that such technological advances be rapidly translated into real-world applications, ensuring that enzymatic PET degradation becomes a cornerstone of sustainable waste management in the coming decades.
Acknowledgments
I cannot express enough gratitude for my mentor who closely guided me throughout this process: Dr. Cesar Carrasco-Lopez. Finally, to everyone around me, like my mother and father for supporting me in this journey of research.
References
- Geneva Environment Network. Plastics and human health | plastics and the environment series. Geneva Environment Network (2022). [↩]
- S. Bertolazzi, A. Cuttitta, V. Pipitone. Addressing marine plastic pollution: a systematic literature review. Current Opinion in Environmental Sustainability 68, 101428 (2024). [↩]
- C. Blanco, C. Spanbauer, S. Stienecker. America’s broken recycling system. California Management Review Insights 66 (2023). [↩] [↩]
- M. G. Kibria, N. I. Masuk, R. Safayet, H. Q. Nguyen, M. Mourshed. Plastic Waste: Challenges and Opportunities to Mitigate Pollution and Effective Management. International Journal of Environmental Research and Public Health 17(1) (2023). [↩]
- Z. Gilani. Enzymatic Depolymerization and Recycling: Using Enzymes to Convert Linear Waste Streams into Circular Supply Chains. Cleantech Group (2022). [↩] [↩]
- G. D. Rocco, H. N. Taunt, M. Berto, H. O. Jackson, D. Piccinini, A. Carletti, G. Scurani, N. Braidi, S. Purton. A PETase enzyme synthesised in the chloroplast of the microalga Chlamydomonas reinhardtii is active against post-consumer plastics. Scientific Reports 13, 10028 (2023). [↩] [↩]
- S. Tanasupawat, T. Takehana, S. Yoshida, K. Hiraga, K. Oda. Ideonella sakaiensis sp. nov., isolated from a microbial consortium that degrades poly(ethylene terephthalate). International Journal of Systematic and Evolutionary Microbiology 66, 2813–2818 (2016). [↩] [↩]
- F. Kawai. The current state of research on PET hydrolyzing enzymes available for biorecycling. Catalysts 11, 206 (2021). [↩] [↩]
- Zero Waste Europe. The impact of Waste-to-Energy incineration on climate. Policy Briefing 2 (2019). [↩]
- E. Doniavi, R. Babazadeh, R. Hasanzadeh. Polyethylene gasification for sustainable plastic waste management with respect to energy, exergy, and environmental considerations: a non-linear programming optimization. Process Safety and Environmental Protection 182, 86–97 (2024). [↩]
- B. Ciuffi, E. Fratini, L. Rosi. Plastic pretreatment: the key for efficient enzymatic and biodegradation processes. Polymer Degradation and Stability (2024). [↩]
- T. S. Ogunlusi, S. S. Ikoyo, M. Dadashipour, H. Gao. Engineering IsPETase and Its Homologues: Advances in Enzyme Discovery and Host Optimisation. International Journal of Molecular Sciences 26, 6797 (2025). [↩] [↩]
- A. L. Aloia, R. V. Glatz, E. J. McMurchie, W. R. Leifert. GPCR expression using baculovirus-infected Sf9 cells. Methods in Molecular Biology 552, 115–129 (2009). [↩] [↩]
- X. Shi, D. L. Jarvis. Protein N-glycosylation in the baculovirus–insect cell system. Current Drug Targets 8(10), 1116–1125, https://doi.org/10.2174/138945007782151315 (2007). [↩]
- H. Chai, S. G. Vasudevan, A. G. Porter, K. L. Chua, S. Oh, M. Yap. Glycosylation and high-level secretion of human tumour necrosis factor-beta in recombinant baculovirus-infected insect cells. Biotechnology and Applied Biochemistry 18(3), 259–273 (1993). [↩]
- B. Deng, Y. Yue, J. Yang, M. Yang, Q. Xing, H. Peng, F. Wang, M. Li, L. Ma, C. Zhai. Improving the activity and thermostability of PETase from Ideonella sakaiensis through modulating its post-translational glycan modification. Communications Biology 6, 1–10 (2023). [↩]
- C. L. Kielkopf, et al. Purification of Polyhistidine-Tagged Proteins by Immobilized Metal Affinity Chromatography. Cold Spring Harbor Protocols 2020(6), pdb.prot102194 (2020). [↩]
- P. G. C. N. T. Pilapitiya, A. S. Ratnayake. The world of plastic waste: a review. Cleaner Materials 11, 100220 (2024). [↩]
- A. L. Aloia, R. V. Glatz, E. J. McMurchie, W. R. Leifert. GPCR expression using baculovirus-infected Sf9 cells. Methods Mol Biol. 552, 115–129 (2009). [↩]
- V. Pirillo, L. Pollegioni, G. Molla. Analytical methods for the investigation of enzyme-catalyzed degradation of polyethylene terephthalate. The FEBS Journal 288, 4730–4745 (2021). [↩] [↩]
- L. L. B. Kastberg, R. Ard, M. K. Jensen, C. T. Workman. Burden imposed by heterologous protein production in two major industrial yeast cell factories: identifying sources and mitigation strategies. Frontiers in Fungal Biology 3 (2022). [↩]
- L. M. Carter, C. E. MacFarlane, S. P. Karlock, T. Sen, J. L. Kaar, J. A. Berberich, J. T. Boock. Increased cytoplasmic expression of PETase enzymes in E. coli. Microbial Cell Factories 23, 319 (2024). [↩]
- R. A. Scheel, T. Ho, Y. Kageyama, J. Masisak, S. McKenney, B. R. Lundgren. Optimizing a fed-batch high-density fermentation process for medium chain-length poly(3-hydroxyalkanoates) in Escherichia coli. Frontiers in Bioengineering and Biotechnology 9, 618259 (2021). [↩]
- C. L. Ward, E. W. Kranenburg. Biological degradation of polyester polyurethane by Comamonas acidovorans. Polymer Degradation and Stability 91(12), 2739–2746 (2006). [↩]
- J. Shiloach, R. Fass. Growing E. coli to high cell density — a historical perspective on method development. Biotechnology Advances 23(5), 345–357 (2005). [↩]
- N. Wang, N. Wang, F. Guan, X. Lv, D. Han, Y. Zhang, N. Wu, X. Xia, J. Tian. Enhancing secretion of polyethylene terephthalate hydrolase PETase in Bacillus subtilis WB600 mediated by the SP amy signal peptide. Letters in Applied Microbiology 71, 235–241 (2020). [↩]


