
Ageing is an inevitable biological process. Unable to resist against the test of time, all normal cells will eventually cease reproduction and break down. In the past, age-related disorders were not major problems, as people did not live long enough to experience growing old. For much of human history, average life expectancies have wavered around 30-40 years old1. Owing to the advent of modern medicine and improved hygiene, the average life expectancy in the United States has now skyrocketed to 77 years1. Because people live longer than they used to, once rare age-related disorders are becoming more prominent in modern society. Thus, an era of anti-ageing medicine has been born to combat growing old, a fairly new phenomenon.
A major pillar in anti-ageing research is figuring out the molecular causes behind getting old. Numerous studies point to reactive oxygen species (free radicals) that accumulate over time and damage the DNA2. Indeed, elevated levels of reactive oxygen species have been found to correlate with ageing and oxidative stress3. Other intracellular factors, such as accelerated telomere shortening due to stress, have also been implicated in the ageing process4.
Like other cells, neurons are not an exception from the ill effects of ageing. Normally, peripheral neurons can regenerate better than spinal cord and brain neurons due to an inhibitory glial environment in the central nervous system5. However, all neurons decline in regenerative capacity over time even in permissive environments. In 1995, Chen and colleagues compared the regenerative capacity of different aged retinal neurons grown in different environments following axonal injury6. Injured embryonic retinal axons were able to regrow even in an older inhibitory glial environment, while postnatal day 2 or older retinal axons failed to regrow even in young permissive environment. Subsequent studies also show that young neurons transplanted in vivo into older spinal cord can regenerate axons, indicating a cell-autonomous mechanism behind neural regenerative failure7. However, the underlying pathway that controls developmental decline of neuronal regeneration remains poorly understood. A new study by Zou and colleagues has elucidated this mystery by identifying a developmental neuron-intrinsic molecular pathway that leads to regenerative failure8. By targeting a step in this pathway, the researchers were able to turn back the clock on old neurons and allow their axons to regenerate extensively again!

Figure 1: A) Caenorhabditis elegans, free living worm widely used to study developmental processes. B) Due to its transparency, different cells can be labeled with fluorescent molecules for visualization and imaging under a microscope. Here, green and yellow fluorescence label a neural circuit involved in C. elegans movement. Images taken from Immortal Humans.com and Pigott et al, 2013.
Caenorhabditis elegans (C. Elegans) is a free-living worm that has been used extensively to study animal development and behavior. The soil nematode offers great potential for scientific study due to its rapid (3-day) life cycle, small size, and ease of laboratory cultivation9). More importantly, C. Elegans is transparent, allowing for in-vivo analysis of the nervous system. Similar to mammals, the worm gets worse at regenerating injured axons as it ages10. Since heterochronic genes (genes that control timing of age-specific events) are important in controlling development of C. Elegans, Zou and colleagues hypothesized that they are also implicated in controlling switches that turn neuronal regeneration on or off. Using C. Elegans as an animal model, the researchers discovered a microRNA signaling pathway that can be manipulated to restore youthful regenerative capacity to old neurons.
MicroRNA (miRNA) is a molecule that is crucial in post-transcriptional gene expression control. Typically a short single stranded nucleotide strand, miRNA require dicer and argonaute proteins for biogenesis11. After it interacts with argonaute and other proteins to form the RNA-induced silencing complex (RISC), miRNA can bind to target mRNA strands, blocking translation of certain genes (Figure 2). This procedure of gene silencing is critical for controlling which genes to turn on or off during different stages of development. To explore the role of miRNA in regulating neuronal regeneration, Zou and others investigated the ability of anterior ventral microtubule (AVM) axons to regenerate in mutants defective of miRNA biogenesis, dcr-1 (dicer) and alg-1 (argonaute)8.
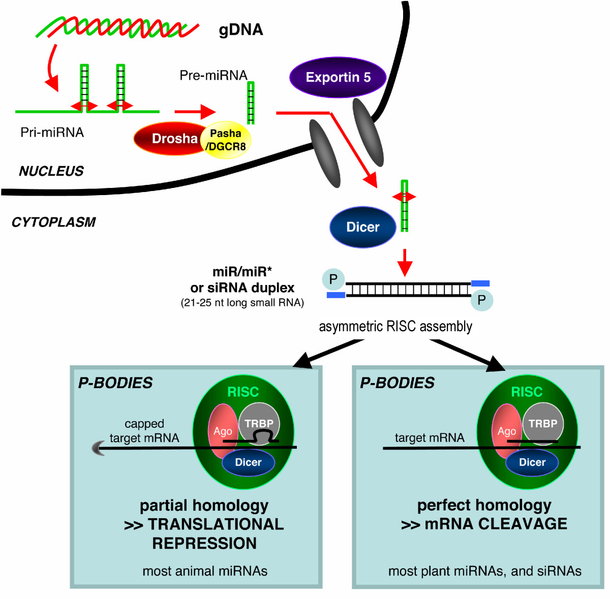
Figure 2: miRNA synthesis and mechanism of action. Dicer cleaves double stranded RNA, giving rise to short single stranded RNA. A short RNA strand then binds to the argonaute protein, forming RISC. The RNA strand then guides RISC to the correct mRNA target, inhibiting translation of certain genes into protein. Image taken from Wikipedia.
In order to induce injury, the researchers used laser to make cuts onto targeted axons. 24 hours after axonal injury at the adult stage, dcr-1 mutants showed no significant improvement in regeneration, while the alg-1 mutants regrew their axons 2.5 times the lengh of those in wild type animals. In addition, adult alg-1 mutants AVM axon shared the same regenerative pattern as younger wild-type axons, indicating that the youthful regenerative capacity was restored in older neurons. To look for miRNAs that interacted with alg-1, a stem-loop reverse transcription polymerase chain rection (RT-PCR) was performed on RNA samples extracted from the animals. Among the 90 miRNAs surveyed, only let-7 was found to be affected by alg-1 and expressed in mature AVM neurons. In addition, let-7 expression was marginal in AVM during larval L1 and L2 stages, but was significantly increased from L3 onward8. This pattern of expression suggests that let-7 is expressed in a timely manner to turn off AVM axonal regeneration at the adult stage.
A well characterized pathway, let-7 is a 21-nucleotide long miRNA that inhibits the lin-41 heterochronic gene12. When lin-41 is not controlled by let-7, it can act to suppress the lin-29 transcription factor13. In larval C. elegans, lin-41 is turned on, suppressing lin-29 to maintain AVM axonal regeneration, while simultaneously inhibiting alg-1 to prevent let-7 miRNA synthesis. As the animal matures, the signaling pathway changes so that let-7 can target lin-41, activating the lin-29 transcription factor to repress regeneration of adult neurons (Figure 3). The target genes of lin-29 that act to inhibit regeneration are currently unknown, which could be an area of worthwhile scientific exploration.
- Machalek, A. (Ed.). (2005). Inside the Cell. [↩] [↩]
- Finkel, T., & Holbrook, N. (2000). Oxidants, oxidative stress and the biology of ageing. Nature. http://dx.doi.org/10.1038/35041687 [↩]
- Tang, S., Le, P., Tse, S., Wallace, D., & Huang, T. (2009). Heterozygous Mutation of Opa1 in Drosophila Shortens Lifespan Mediated through Increased Reactive Oxygen Species Production. PLOS One [↩]
- Epel, E., Blackburn, E., Lin, J., Dhabhar, F., Adler, N., Morrow, J., & Cawthon, R. (2004). Accelerated telomere shortening in response to life stress. Proceedings of the National Academy of Sciences. [↩]
- Yiu, G., & He, Z. (2006). Glial inhibition of CNS axon regeneration. Nature Reviews Neuroscience. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/16858390 [↩]
- Chen, D., Jhaveri, S., & Schneider, G. (1995). Intrinsic changes in developing retinal neurons result in regenerative failure of their axons. Proceedings of the National Academy of Sciences. [↩]
- Blackmore, M., & Letourneau, P. (2006). Changes within maturing neurons limit axonal regeneration in the developing spinal cord. Journal of Neurobiology [↩]
- Zou, Y., Chiu, H., Ambros, V., Zinovyeva, A., & Chang, C. (2013). Developmental Decline in Neuronal Regeneration by the Progressive Change of Two Intrinsic Timers. Science, 372-376. [↩] [↩] [↩]
- Riddle, D., Blumenthal, T., Meyer, B., & Priess, J. (1997). Introduction to C. Elegans. In C. Elegans II. New York, NY: Cold Spring Harbor Laboratory Press. (Original work published 1997 [↩]
- Wu, Z., Ghosh-Roy, A., Yanik, M., Zhang, J., Jin, Y., & Chisolm, A. (2007). Caenorhabditis elegans neuronal regeneration is influenced by life stage, ephrin signaling, and synaptic branching. Proceedings of the National Academy of Sciences. [↩]
- Nix, P., & Bastiani, M. (2013). Heterochronic Genes Turn Back the Clock in Old Neurons. Science, 282. [↩]
- Reinhart, B., Slack, Basson, M., Pasquinelli, A., Bettinger, J., Rougvie, A., . . . Ruvkun, G. (2000). The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature. [↩]
- Slack, Basson, Liu, Ambros, Horvitz, & Ruvkun. (2000). The lin-41 RBCC gene acts in the C. elegans heterochronic pathway between the let-7 regulatory RNA and the LIN-29 transcription factor. Cell. [↩]
- Chen, Lai, & Niswander. (2012). The ubiquitin ligase mLin41 temporally promotes neural progenitor cell maintenance through FGF signaling. Genes and Development [↩]



[…] The sample article to suggest from this publication is: Going back in time with the microRNA neural time machine: from old neurons to young neurons. It is in this link: https://nhsjs.com/2013/going-back-in-time-with-the-microrna-neural-time-machine-from-old-neurons-to-y… […]