
Emily Bo1, Qing Song2 & Kai Yuan2,3,4,5
1 Lincoln High School, Seattle, Washington, United States.
2 School of Life Science and Technology, Xidian University, Xi’an, Shaanxi 710126, China.
3 Inner Mongolia Key Laboratory of Pattern Recognition and Intelligent Image Processing, School of Information Engineering, Inner Mongolia University of Science and Technology, Baotou, Inner Mongolia 014010, China.
4 Xi’an Key Laboratory of Intelligent Sensing and Regulation of trans-Scale Life Information, School of Life Science and Technology, Xidian University, Xi’an, Shaanxi 710126, China.
5 Engineering Research Center of Molecular and Neuro Imaging Ministry of Education, Xi’an, Shaanxi 710126, China.
Abstract
Background and Hypothesis: Excessive and uncontrolled gaming behavior in those with Internet Gaming Disorder is associated with impulsivity, which may correlate with reward network and cerebellar disruptions. Recent studies have uncovered the cerebellum’s role in regulating impulsive activity. We hypothesized that decreased connectivity between the reward network and cerebellar regions would be found in IGD patients and that impulsivity would be negatively correlated with functional connectivity.
Methods: This study used resting-state functional magnetic resonance imaging (fMRI) to investigate whether differences in functional connectivity exist in adolescents with IGD. Resting-state fMRI (rsFMRI) data of 89 adolescents with IGD and 101 healthy controls was analyzed using network-based analysis (NBS) to identify abnormal subnetworks, as well as exploratory seed-based analysis to identify specific abnormal connections. Spearman’s correlation analysis was conducted between Barratt Impulsiveness Scale-11 (BIS-11) scores and in IGD patients.
Results: Network-based analysis (NBS) revealed one disrupted subnetwork in IGD, with major disruptions between the thalamus, hippocampus, and cerebellum (lobules VI, V, and III). Four additional subnetworks were significantly associated with impulsivity, with disruptions between the accumbens and lobule VIIA (Crus I and II). Seed-based analysis (SBA) showed decreased connectivity with multiple cerebellar lobules (Crus I/II, lobule VIII/IX). Correlation analysis showed a negative correlation between functional connectivity and impulsivity.
Conclusion: Our results highlight cerebellum-reward network connectivity disruptions in those with IGD and in the development of increased impulsivity, which could be a possible target for future intervention.
Introduction
Video games have skyrocketed in popularity in the past 20 years, and with it rates of problematic gaming have risen as well. A 2018 meta-analysis calculated the prevalence of IGD in adolescents as 4.6%1, and many more find gaming a daily occurrence.
Increased impulsivity is often observed in patients with IGD2, which in turn may be associated impaired self-regulation and excessive gaming behavior3. Impulsivity is also positively associated with reward network dysfunction, particularly in circuits involving the striatum, orbitofrontal cortex (OFC), and anterior prefrontal regions4. Within this network, the hippocampus contributes to tracking motivational salience5, the OFC encodes reward value6, and regions of the striatum support reward prediction and error7,8. Disruptions in this network are hypothesized to promote behavior favoring immediate rewards and weaken regulatory control, contributing to impulsive decision making4.
Though traditionally associated with motor functions, recent evidence supports the cerebellum’s role in cognition, risk tolerance, and impulsivity9,10,11.
Subcortical‐cerebellar connectivity is correlated with self‐reported impulsivity4. Lower striato-cerebellar functional connectivity strength is associated with a higher tendency to avoid or inhibit actions with possibly negative outcomes such as punishment, a characteristic of impulsivity12. In addition, the cerebellum plays a role in inhibiting circuits responsible for initiating actions, as animal and human patients with cerebellar lesions exhibited impaired behavioral inhibition9. The cerebellum connects functionally to the striatum13, a component of the reward network14.
The pathophysiology of IGD, as well as many other neuropsychiatric disorders, implicates the cerebellum. Neuromodulation studies suggest that stimulating cerebellar regions can alter cortical activity15, which may reduce impulsive behaviors9, especially in IGD patients. When distinguishing healthy controls from IGD patients, both the anterior and posterior lobes of the cerebellum were the most impacted16. In another study, using multi-variate pattern analysis to analyze rsfmri data, the bilateral inferior cerebellum displayed the strongest discriminative power when classifying IGD17. Given previous evidence suggesting a role of the cerebellum in IGD, we investigated whether cerebellar interactions with the reward network are altered in IGD. This question provides the rationale for the current study.
Literature Review
Previous studies on cerebellar connectivity in Internet Gaming Disorder (IGD) have primarily examined connectivity between the cerebellum and the entire brain18, rather than connectivity between the cerebellum and individual regions in the brain. In addition, investigations focusing specifically on altered connectivity between the cerebellum and the reward network remain limited, so we sought to investigate this connectivity. In the present study, we analyzed resting-state functional connectivity data using predefined regions of interest (ROIs) within the cerebellum and reward network to identify abnormal networks and connectivity patterns and their associations with impulsivity.
Based on prior research on cerebellar-reward network interactions19, we hypothesized that individuals with IGD would exhibit decreased functional connectivity between the cerebellum and reward-related regions, correlating with greater impulsivity severity. By elucidating the neural networks and pathways implicated in impulsivity within IGD, this study aims to determine whether alterations in cerebellar connectivity are associated with IGD.
Methods
Participants
This study utilized data from two scanning sites. Scanning site 1 consisted of 34 IGD and 41 HC participants from Xiangya, China. Scanning site 2 consisted of 55 IGD and 60 HC participants from Shanghai, China. Scanning site 1 contained both male and female participants, while scanning site 2 contained only male participants. Data from these sites, which contains differing sex compositions, were pooled to model sex-related effects.
All participants were Chinese and all IGD participants met the diagnostic criteria according to International Classification of Diseases 11th Revision (ICD-11). No psychiatric comorbidities and other neurological disorders were present in both healthy and IGD participants and no participants used medications. Demographic information for both sites is provided in Table 1. A one-tailed priori power analysis using G*Power20 indicated that 122 participants were required to detect a medium effect size (r = 0.30) with 80% power at  = 0.005.
= 0.005.
Abbreviations: HC, healthy controls; IGD = patients with Internet Gaming Disorder; BIS = Barratt Impulsiveness Scale; SD = standard deviation.
Measures
Impulsivity was measured using the 11th version of the Barett Impulsivity Scale(BIS-11)21.
Image Acquisition
For both sites, MRI data was acquired using a 3-T scanner with a standard head coil. Before the scanning, all subjects were instructed to keep their eyes closed, relax, keep their head very still and think of nothing. Foam pads were used in both sites to minimize head motion and reduce scanner noise.
For Site 1, participants were scanned using a Siemens Skyra 3-T Scanner. The resting-state BOLD fMRI images were obtained by a single-shot, gradient-recalled echo-planar imaging sequence with the following scanning parameters: repetition time = 2,000 ms; echo time = 30 ms; slice gap = 1 mm; matrix size = 64 x 64; flip angle = 80°; field of view = 240 x 240 mm; slice thickness = 4 mm and number of slices = 32. For each participant, 216 volumes were obtained, and the scan lasted 432 s.
For Site 2, participants were scanned using a 3-T GE Signa HDx Scanner. The resting-state BOLD fMRI images were obtained by a single-shot, gradient-recalled echo-planar imaging sequence with the following scanning parameters: repetition time = 2,000 ms; echo time = 24 ms; no slice gap; matrix size = 64 x 64; flip angle: 90°; field of view = 230 x 230 mm; slice thickness = 4 mm; number of slices = 34. For each participant, 220 volumes were obtained, and the scan lasted 440 s.
Image Preprocessing
For both sites, structural and functional MRI data were preprocessed using AFNI22,23 (version 25.2.06) and FSL24 (version 5.0.1). T1-weighted anatomical images were deobliqued, reoriented to Right–Posterior–Inferior (RPI) orientation, skull-stripped, and a binary brain mask was generated and applied using voxel-wise multiplication. For functional images, the first 10 volumes were excluded from analysis. Then, functional images were deobliqued, reorientated to RPI orientation, and motion corrected using Fourier interpolation, two-pass registration, and z-padding (4). Then, the images were skull-stripped, followed by spatial smoothing (4 mm FWHM), intensity normalization (10,000), temporal bandpass filtering (0.009–0.1 Hz) and polynomial detrending.
Anatomical images were segmented into gray matter, white matter (WM), and cerebrospinal fluid (CSF) masks using FSL’s FAST. WM and CSF masks were transformed into functional space, smoothed, and refined using tissue priors in the MNI space. The resulting masks were thresholded and intersected with the functional brain mask.
Functional-to-anatomical registration was performed using rigid-body (6 DOF) linear alignment with a correlation ratio cost function. Anatomical-to-standard (MNI152 2 mm) registration used affine (12 DOF) linear alignment with correlation ratio cost. The functional-to-standard registration was obtained by concatenating both transformations and was resampled into standard space using the concatenated matrix. All transformations utilized FSL’s FLIRT with trilinear interpolation.
Nuisance regression was performed on preprocessed functional data using a voxelwise GLM using FSL’s FILM. The model included six rigid-body motion parameters, mean signals extracted from the whole brain, CSF, and WM masks derived from subject-specific segmentation and aligned to functional space. The WM and CSF regressors were included to reduce non-neuronal variance through physiological noise or scanner drift. Motion regressors were included to mitigate motion-related artifacts and distance-dependent bias. The mean global signal was included as a regressor to reduce widespread physiological/vascular noise and motion-related global artifacts and did not materially affect correlation structure.
A design matrix was generated using a FEAT-based template with TR = 2 s and 221 timepoints. Next, regression was performed using FILM with prewhitening and spatial autocorrelation smoothing (5). The minimum voxel intensity within the brain mask was used as the threshold for model fitting. Residual time series were extracted, mean-centered, and shifted by a constant (+100) to preserve positive intensity values via this equation:
![\[ residual = (a - \bar{a}) + 100\]](https://nhsjs.com/wp-content/ql-cache/quicklatex.com-1f9f9696d88ab68af0e33f61b713d24d_l3.png "Rendered by QuickLaTeX.com")
Network-Based Analysis
We chose 44 ROIs (18 reward network ROIs and 26 cerebellar ROIs). For reward network ROIs, we selected regions based on Harber and Knutson’s proposed model of the reward network14. For the cerebellar ROIs, we selected regions based on the Automated Anatomical Labelling Atlas. We extracted each reward network ROI individually from the Harvard-Oxford Subcortical and Harvard-Oxford Cortical Atlas and each cerebellar ROI from the Automated Anatomical Labelling Atlas25 via FSL. Then, we combined each individual ROI into one atlas file, extracting the mean signal from each ROI using DPARSFA26 to create a 44 x 44 functional connectivity matrix and Fisher-Z transformed to enhance normality.
Group-level comparisons were performed using a Generalized Linear Model (GLM) in CONN v204727, controlling for age, gender, and site. Network-based statistics28 identified abnormal subnetworks through non-parametric inference and grouped related ROI connections into subnetworks. Abnormal subnetworks were identified with a connection threshold of P < 0.001, followed by network-level significance testing with False Discovery Rate (FDR) correction (pFDR-corrected < 0.05). The same methods and thresholds were applied to identify networks responsible for impulsivity in those with IGD, controlling for age, gender and scan site.
Seed-Based Analysis
Seed-based analyses were conducted as exploratory follow-up analyses. As the functional organization of the cerebellum does not always align with anatomical subdivisions, we selected network regions from the subnetwork identified via NBS to determine the specific cerebellar coordinates with altered connectivity. To avoid data reuse bias (double-dipping), we are unable to draw conclusions regarding seed-based analysis results.
The mean signal from these reward network regions was extracted and correlated with whole-brain voxels using Pearson correlation, followed by Fisher Z transformation to enhance normality. Group-level functional connectivity differences between IGD patients and HCs were then assessed using SPM’s GLM, controlling for age, gender, and site, with statistical significance set at cluster-level pFDR-corrected < 0.05 and voxel level P < .001.
Correlation analysis between functional connectivity and symptoms
We examined the relationship between abnormal functional connectivity and impulsivity in patients with IGD using Spearman’s rank correlation, controlling for age, gender, and site and applying the Benjamini–Hochberg FDR procedure (q < 0.05) for multiple comparisons29. We specifically focused on abnormal networks found via network-based analysis.
Results
Network-Based Analysis
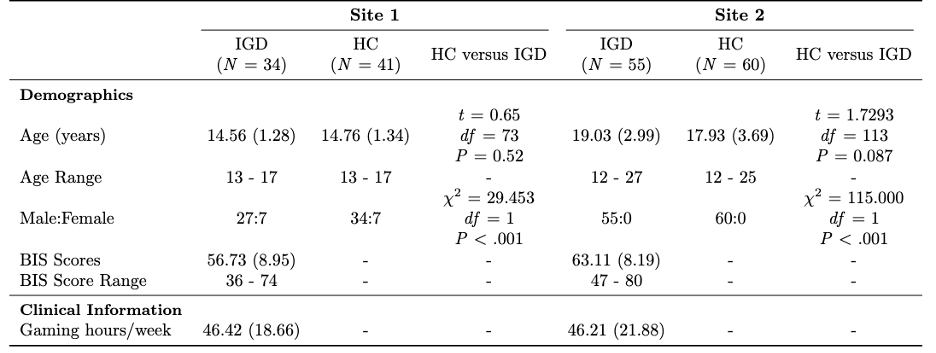
Network-Based Analysis found 1 disrupted subnetwork reflecting reduced connectivity in IGD relative to HC, containing 210 connections among 21 ROIs (Figure 1a). This subnetwork primarily had disrupted connectivity between the thalamus, hippocampus, and the cerebellum, as well as within the cerebellum (Network Mass = 1702.49, pFDR < 0.001).
Network-based analysis also identified 4 significant subnetworks associated with BIS scores in patients with IGD (Figure 1b). These networks involved disrupted cerebellar connections with thalamic and striatal (accumbens and putamen), as well as intra-cerebellar connectivity. “Subnetwork 1” consisted of 6 connections and primarily involved abnormal connectivity between the thalamus, hippocampus, vermis (vermis III, Network Mass = 166.57, pFDR < 0.001). “Subnetwork 2” comprised 4 connections between the accumbens, putamen and vermis (vermis IV-V, Network Mass = 105.77, pFDR < 0.001). “Subnetwork 3” contained 1 connection between the right middle frontal gyrus and the right cerebellum crus (Network Mass = 24.46, pFDR = 0.02). “Subnetwork 4” contained 1 connection between the right Crus I and left Crus II (Network Mass = 23.67, pFDR = 0.021).
Seed-Based Analysis
We used the significant regions identified in network-based analysis, that being the thalamus, hippocampus, caudate, and putamen, to conduct seed-to-voxel analysis.
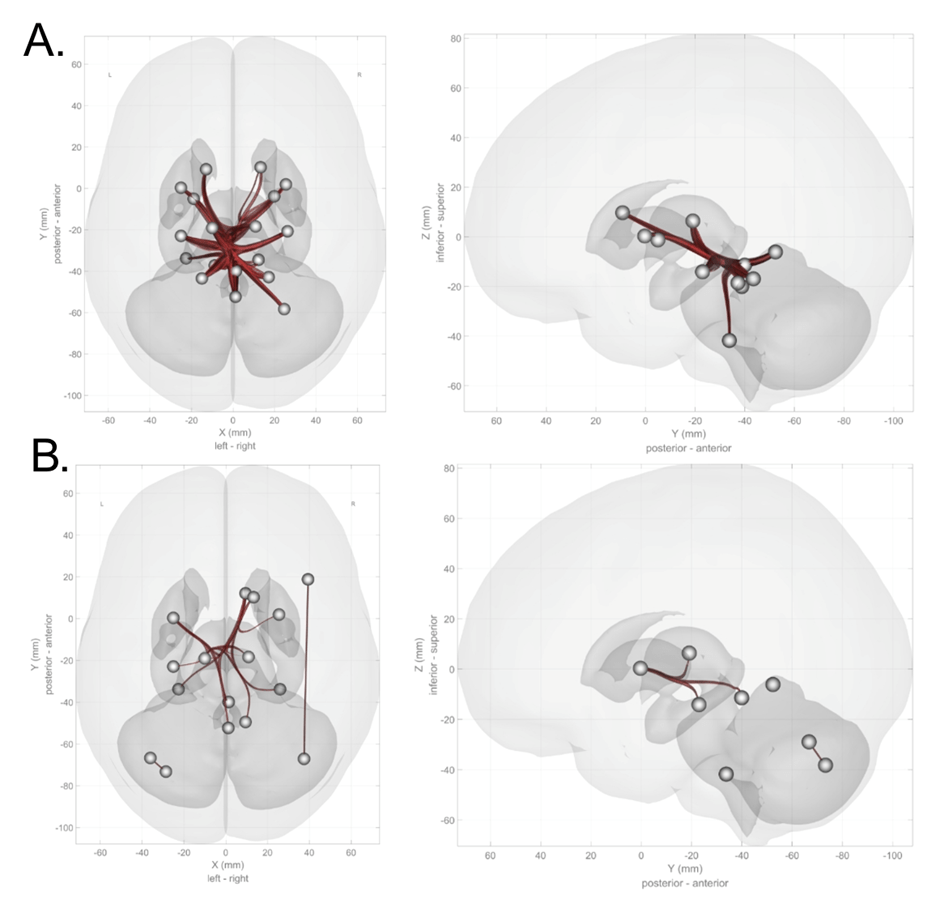
For the left hippocampus, seed-based analysis discovered a significant decrease in connectivity with the left cerebellum crus I (Figure 2; peak coordinates: (-38, -56, -52), t = 4.8; cluster-level pFDR-corrected < 0.001; voxel-level P < .001; cluster size = 347 voxels). For the right hippocampus, seed-based analysis discovered a significant decrease in connectivity with the left cerebellum lobule VIII (peak coordinates: (-36, -58, -46), t = 5.9; cluster-level pFDR-corrected < 0.001; voxel-level P < .001; cluster size = 492 voxels) as well as in the right cerebellum crus I (peak coordinates: (18, -74, -30), t = 5.3; cluster-level pFDR-corrected = 0.03; voxel-level P < .001; cluster size = 77 voxels).
For the left putamen, a significant decrease was found in connectivity with the left cerebellum crus II (peak coordinates: (-08, -72, -34), t = 5.4; cluster-level pFDR-corrected < 0.001; voxel-level P < .001; cluster size = 171 voxels). For the right putamen, seed-based analysis discovered a significant decrease in connectivity with the right cerebellum lobule IX (peak coordinates: (+10 -42 -42), t = 5.7; cluster-level pFDR-corrected = 0.003; voxel-level P < .00; cluster size = 367 voxels).
Interestingly, for the left caudate, we found a significant increase in functional connectivity with the right cerebellar lobule VI (peak coordinates: (16, -74, -24), t = 8.7; cluster-level, pFDR-corrected < 0.001; voxel-level P < .001; cluster size = 788 voxels). For the right caudate, we also found an increase in functional connectivity, this time with the left cerebellum crus I (peak coordinates: (-38, -66, -44), t = 5.6; cluster-level, pFDR-corrected < 0.001; voxel-level P < .001; cluster size = 804 voxels).
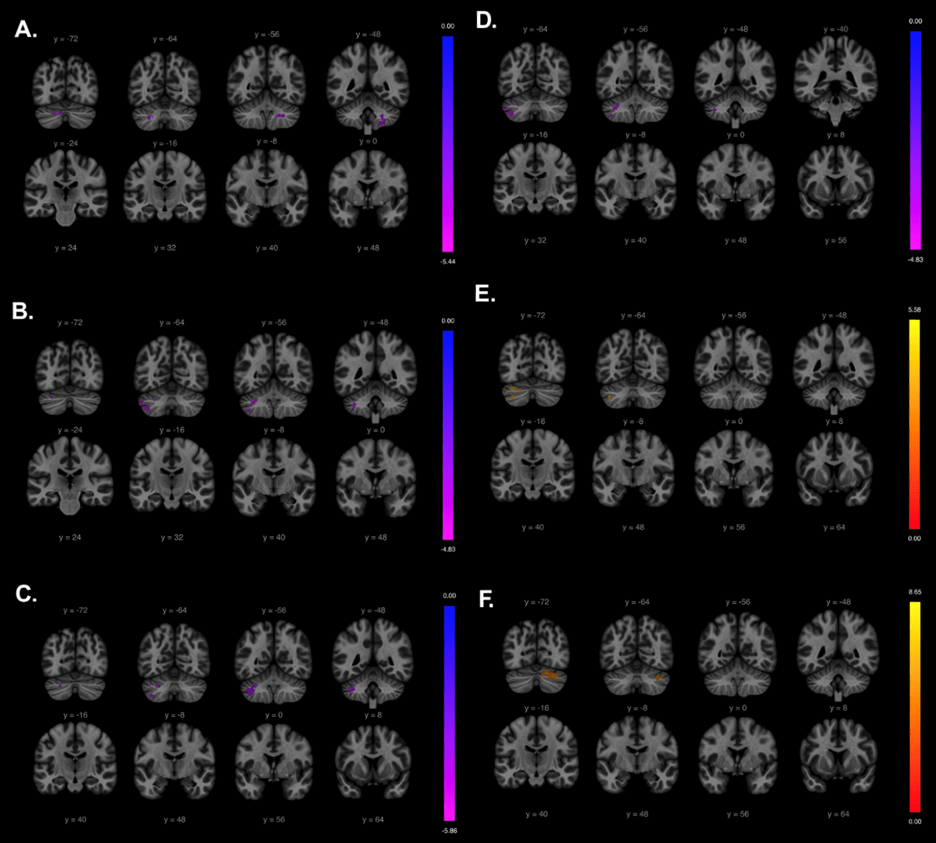
The thalamus yielded no significant results during seed-to-voxel analysis.
Correlation Analysis Between Functional Connectivity and Symptoms
IGD patients exhibited significantly reduced functional connectivity of the left cerebellar lobule VI with both the putamen and caudate nucleus. We also sought to examine whether there would be a positive correlation with impulsivity when assessing connectivity between the caudate and the cerebellum. We conducted 4 correlation analyses between connectivity strength (lobule V–hippocampus, lobule X–thalamus, lobule III–thalamus) and impulsivity (Figure 3). These 4 connections were chosen as they had the highest network mass out of all connections when conducting network-based analysis. We controlled for sex, age, and site. Connectivity between the right lobule V and the left hippocampus was significantly negatively correlated with impulsivity (Spearman’s  = -0.435, P <0.001). Lobule X–left thalamus also showed negative correlation with impulsivity (Spearman’s = -0.279, P = .008). Right lobule III–left thalamus connectivity showed significant negative correlations with impulsivity (Spearman’s = −0.222, P = .037). The Crus I–caudate connection, however, was positively correlated with BIS scores (Spearman’s = 0.298, P = .005). All four connections remained significant after FDR correction (q < 0.05).
= -0.435, P <0.001). Lobule X–left thalamus also showed negative correlation with impulsivity (Spearman’s = -0.279, P = .008). Right lobule III–left thalamus connectivity showed significant negative correlations with impulsivity (Spearman’s = −0.222, P = .037). The Crus I–caudate connection, however, was positively correlated with BIS scores (Spearman’s = 0.298, P = .005). All four connections remained significant after FDR correction (q < 0.05).
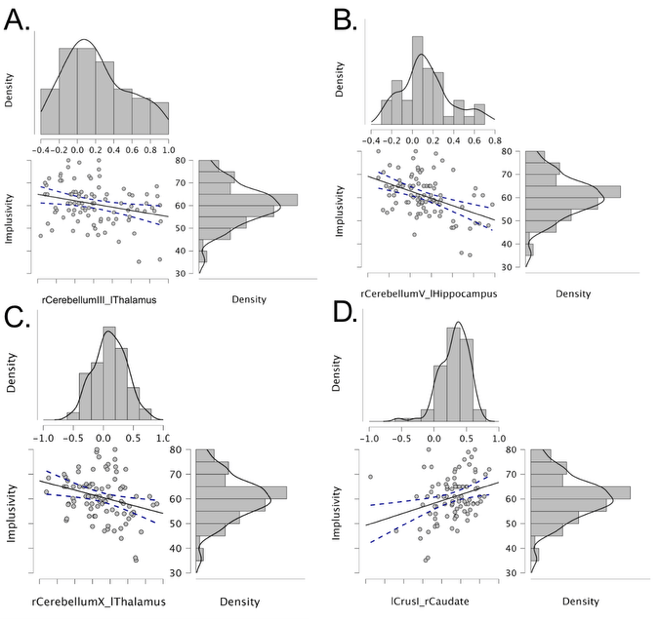 = −0.222, P = .037) (B) Negative correlation between cerebellar lobule V–hippocampus (Spearman’s = −0.435, P <0.001) (C) Negative correlation between cerebellar lobule X–thalamus (Spearman’s = −0.279, P = .008) (D) Positive correlation between Crus I–caudate and impulsivity. (Spearman’s = 0.298, P = 0.005)
= −0.222, P = .037) (B) Negative correlation between cerebellar lobule V–hippocampus (Spearman’s = −0.435, P <0.001) (C) Negative correlation between cerebellar lobule X–thalamus (Spearman’s = −0.279, P = .008) (D) Positive correlation between Crus I–caudate and impulsivity. (Spearman’s = 0.298, P = 0.005)Discussion
This study examined differences in functional connectivity between the cerebellum and the reward network in IGD. Network-based analysis revealed abnormalities in the cerebellar–basal ganglia network, with the most pronounced abnormalities in the thalamus, caudate, hippocampus, and putamen. Reduced functional connectivity in the accumbens and separately, in the middle frontal gyrus, was also observed when associating functional connectivity with BIS scores.
The thalamus, caudate, hippocampus, and putamen are key components of the basal ganglia, a collection of anatomically and functionally distinct subcortical regions responsible not only for motor control, but also reward processing and coding prediction error.30,31. The thalamus provides an alerting signal that shapes striatal action selection when anticipating reward32. Decreased connectivity between the thalamus and the cerebellum could be associated with altered integration of reward, timing, and cognitive control processes, which also involve the thalamus.
The hippocampus, which is involved in memory encoding33, has also been implicated in IGD. Structural or functional alterations in this region may be linked to greater preference for immediate rewards34. The putamen and caudate comprise the dorsal striatum. In addition to maintaining information about the rewarding outcomes of actions and selecting and initiating actions based on rewards35, it also receives dopaminergic projections from the substantial nigra36. The decreased functional connectivity found in both regions could reflect an abnormality in action initiation based on reward. A previous meta-analysis found reduced functional connectivity in the striatum for those with IGD across 30 studies37, and a separate study found decreased connectivity in the putamen38. Our finding of altered connectivity in the accumbens in relation to impulsivity aligns with current evidence that different regions of the nucleus accumbens play regulate distinct facets of impulsive behavior39. This suggests that disruptions in functional connectivity within these regions is associated with increased impulsivity and greater illness severity, consistent with prior reports of a positive correlation between impulsivity and IGD severity40,41.
Abnormalities in these previously identified regions have been reported in those with IGD42. An fMRI study on caudate, putamen, and nucleus accumbens volumes found increased volume in those with IGD and that caudate volume correlated with Stroop task performance43. These findings are consistent with the altered connectivity in found in those areas in the present study. However, results in previous literature have been mixed, with one task-based fMRI study finding increased activation in the left Crus II44 and another finding decreased activation in the left Crus I45.
We identified cerebellar lobules V, Crus I, and Crus II as regions with the most significant decreased connectivity with the reward network. Crus I and II are located in the posterior cerebellum, a region implicated in emotional processing46. Meta-analyses have suggested that the posterior lobe, particularly lobules VI and Crus I, is involved in working memory, spatial tasks, executive functions, and emotional processing47. Previous studies have reported a reduction in gray matter volume in the Crus I and II lobules in individuals with IGD48, with these differences associated with cognitive alteration and increased impulsivity. Animal studies demonstrated interconnected activity between the cerebellum and basal ganglia49, while human research has reported similar results. Negative connectivity between the cerebellum and components of the reward network has been reported in other substance use disorders, like mehampetamine50 and alcohol use disorder51,52. The negative connectivity in individuals with IGD may reflect patterns that overlap with those reported in other substance use disorders.
Correlation analyses between BIS impulsivity scores and connectivity strength in the cerebellum found a strong negative correlation, especially in cerebello–striatal and cerebello–mesolimbic networks. Notably, connectivity strength across several cerebellar–subcortical pathways (lobule V–hippocampus, lobule X–thalamus, and lobule III–thalamus) showed robust negative correlations with impulsivity, even after controlling for demographic and site-related factors. Higher BIS scores were associated with weaker connectivity within these circuits. Prior studies have found that blunted cerebellar influence on reward and memory-related subcortical structures is associated with heightened impulsivity53 in IGD. A previous study on 1,245 first- and second-year college students found that impulsivity greatly predicted tendencies for IGD, and that the strength of thalamic-cortical connectivity also helped predict IGD tendency54. Taken together, our findings suggest that disrupted functional connectivity between the cerebellum and basal ganglia is associated with impulsivity and may represent a neural correlate of maladaptive reward-related behavior in IGD.
Our results indicate that altered cerebellar connectivity is associated with impulsivity and maladaptive decision-making in IGD and highlights cerebellar–reward network connectivity for further investigation in therapeutic research. Prior studies have demonstrated that non-invasive cerebellar stimulation is associated with improved impulsivity in individuals with IGD55, indicating the need for additional study on the modulation of cerebellar circuits. Our research suggests that targeting specific lobules, like the Crus I and II, may be particularly relevant to target impulsivity-related symptoms, though further replication and validation is necessary. Previous studies have found the cerebellum to be associated with other symptoms of IGD, like adehonia56. Thus, future research should further examine the impact of substance use disorders on cerebellar structure and function, as well as the role of cerebellar–reward network connectivity across substance use disorders.
Limitations
The samples of study only included East Asian, specifically, Han Chinese, individuals. The culture around gaming is different in East Asia compared to the rest of the world, and many IGD studies have focused on Asian populations where the diagnosis is more commonplace57. Future studies should include a diverse group of adolescents for study and should recruit older individuals as well.
Scanner- or site-related variability was introduced due to multi-site acquisition of data despite statistical control for site effects. In addition, the sex composition of our samples were not balanced, with one site having few female-identifying participants and the other site having no female-identifying participants, limiting generalizability and potentially confounding gender and site-related differences. Self-report measures used to report impulsivity are also subject to bias and does not fully capture all aspects of impulsivity. Future studies should incorporate consistent acquisition of fMRI data, gender-balanced samples, and objective behavioral assessments to strengthen the interpretability of these findings.
The cross-sectional design of this study also limits our ability to draw causal inferences regarding the observed connectivity alterations. Specifically, it remains unclear whether disrupted cerebellar–striatal coupling reflects a predisposed neural vulnerability for IGD in certain patient groups or a consequence of prolonged excessive gaming. Future longitudinal studies, or investigations tracking individuals during abstinence or treatment, will be necessary to determine whether these connectivity patterns normalize with reduced gaming and recovery.
Conclusion
Disrupted cerebellum–striatum connectivity is associated with IGD and with reduced functional connectivity between thalamus and caudate and posterior cerebellar lobules Crus I and II. Cerebellum–thalamus connectivity is also associated with impulsivity. These findings uncover specific circuit-level associations that could have therapeutic applications but that warrant further validation through hypothesis-gathering experiments.
Acknowledgements
Emily Bo would like to thank Professor Kai Yuan for this amazing summer internship opportunity, Qing Song for support in conducting research, and Rouwan Li for non-research support.
References
- J. Y. Fam. Prevalence of internet gaming disorder in adolescents: A meta-analysis across three decades. Scandinavian Journal of Psychology. Vol. 59, pg. 524–531, 2018, https://doi.org/10.1111/sjop.12459. [↩]
- Y. Zheng, J. He, L. Fan, Y. Qiu. Reduction of symptom after a combined behavioral intervention for reward sensitivity and rash impulsiveness in internet gaming disorder: a comparative study. Journal of Psychiatric Research. Vol. 153, pg. 159–166, 2022, https://doi.org/10.1016/j.jpsychires.2022.06.056. [↩]
- H. Li, O. Turel, Q. He. Neural basis of altered impulsivity in individuals with internet gaming disorder: state-of-the-art review. Current Addiction Reports. Vol. 10, pg. 107–121, 2023 https://doi.org/10.1007/s40429-023-00481-8. [↩]
- P. Hüpen, H. Kumar, D. Müller, R. Swaminathan, U. Habel, C. Weidler. Functional brain network of trait impulsivity: whole‐brain functional connectivity predicts self‐reported impulsivity. Human Brain Mapping. Vol. 45, 2024, https://doi.org/10.1002/hbm.70059. [↩] [↩] [↩]
- M. Sosa, L. M. Giocomo. Navigating for reward. Nature Reviews Neuroscience. Vol. 22, pg. 472–487, 2021, https://doi.org/10.1038/s41583-021-00479-z. [↩]
- E. T. Rolls, W. Cheng, J. Feng. The orbitofrontal cortex: reward, emotion, and depression. Brain Communications. Vol. 2, 2020, https://doi.org/10.1093/braincomms/fcaa196. [↩]
- K. Oyama, I. Hernadi, T. Iijima, K.-I. Tsutsui. Reward prediction error coding in dorsal striatal neurons. Journal of Neuroscience. Vol. 30, pg. 11447–11457, 2010, https://doi.org/10.1523/jneurosci.1719-10.2010. [↩]
- W. Schultz. Reward functions of the basal ganglia. Journal of neural transmission ((Vienna, Austria: 1996), Vol. 123, pg. 679–693, 2016, https://doi.org/10.1007/s00702-016-1510-0. [↩]
- M. Miquel, S. M. Nicola, I. Gil-Miravet, J. Guarque-Chabrera, A. Sanchez-Hernandez. A working hypothesis for the role of the cerebellum in impulsivity and compulsivity. Frontiers in Behavioral Neuroscience. Vol. 13, 2019, https://doi.org/10.3389/fnbeh.2019.00099. [↩] [↩] [↩]
- J. D. Schmahmann. The cerebellum and cognition. Neuroscience letters. Vol. 688, pg. 62–75, 2019, [↩]
- P. Quan, L. He, T. Mao, Z. Fang, Y. Deng, Y. Pan, X. Zhang, K. Zhao, H. Lei, J. A. Detre, J. W. Kable, H. Rao. Cerebellum anatomy predicts individual risk-taking behavior and risk tolerance. NeuroImage. Vol. 254, pg. 119148, 2022, https://doi.org/10.1016/j.neuroimage.2022.119148. [↩]
- M. Abdallah, N. Farrugia, V. Chirokoff, S. Chanraud. Static and dynamic aspects of cerebro-cerebellar functional connectivity are associated with self-reported measures of impulsivity: A resting-state fMRI study. Network neuroscience (Cambridge, Mass.), Vol. 4, pg. 891–909, 2020. https://doi.org/10.1162/netn_a_00149. [↩]
- A. C. Bostan, P. L. Strick. The basal ganglia and the cerebellum: nodes in an integrated network. Nature Reviews Neuroscience. Vol. 19, pg. 338–350, 2018, https://doi.org/10.1038/s41583-018-0002-7. [↩]
- S. N. Haber, B. Knutson. The reward circuit: linking primate anatomy and human imaging. Neuropsychopharmacology. Vol. 35, pg. 4–26, 2010, https://doi.org/10.1038/npp.2009.129. [↩] [↩]
- E. P. Casula, M. C. Pellicciari, V. Ponzo, M. Stampanoni Bassi, D. Veniero, C. Caltagirone, G. Koch. Cerebellar theta burst stimulation modulates the neural activity of interconnected parietal and motor areas. Scientific Reports. Vol. 6, 2016, https://doi.org/10.1038/srep36191. [↩]
- Z.-L. Wang, M. N. Potenza, K.-R. Song, X.-Y. Fang, L. Liu, S.-S. Ma, C.-C. Xia, J. Lan, Y.-W. Yao, J.-T. Zhang. Neural classification of internet gaming disorder and prediction of treatment response using a cue-reactivity fmri task in young men. Journal of Psychiatric Research. Vol. 145, pg. 309–316, 2022, https://doi.org/10.1016/j.jpsychires.2020.11.014. [↩]
- G. Dong, Z. Wang, H. Dong, M. Wang, Y. Zheng, S. Ye, J. Zhang, M. N. Potenza. More stringent criteria are needed for diagnosing internet gaming disorder: evidence from regional brain features and whole-brain functional connectivity multivariate pattern analyses. Journal of Behavioral Addictions. Vol. 9, pg. 642–653, 2020, https://doi.org/10.1556/2006.2020.00065. [↩]
- A. Mundorf, A. Siebert, J. E. Desmond, J. Peterburs. The role of the cerebellum in internet gaming disorder – A systematic review. Addiction biology. Vol 28, pg. e13331, 2023, https://doi.org/10.1111/adb.13331. [↩]
- J. Yoshida, M. Oñate, L. Khatami, J. Vera, F. Nadim, K. Khodakhah. Cerebellar contributions to the basal ganglia influence motor coordination, reward processing, and movement vigor. Journal of Neuroscience. Vol. 42, pg. 8406–8415, 2022, https://doi.org/10.1523/JNEUROSCI.1535-22.2022. [↩]
- F. Faul, E. Erdfelder, A.-G. Lang, A. Buchner. G*Power 3: a flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods. Vol. 51, pg. 175-191, 2007, https://doi.org/https://doi.org/10.3758/bf03193146. [↩]
- J. H. Patton, M. S. Stanford, E. S. Barratt. Factor structure of the barratt impulsiveness scale. Journal of Clinical Psychology. Vol. 51, pg. 768–774, 1995, https://doi.org/https://doi.org/10.1002/1097-4679(199511)51:6<768::AID-JCLP2270510607>3.0.CO;2-1. [↩]
- R. W. Cox. AFNI: software for analysis and visualization of functional magnetic resonance neuroimages. Computers and Biomedical Research. Vol. 29, pg. 162–173, 1996, https://doi.org/10.1006/cbmr.1996.0014. [↩]
- R. W. Cox, J. S. Hyde. Software tools for analysis and visualization of fmri data. NMR in Biomedicine. Vol. 10, pg. 171–178, 1997, https://doi.org/10.1002/(sici)1099-1492(199706/08)10:4/5%3C171::aid-nbm453%3E3.0.co;2-l. [↩]
- S. M. Smith, M. Jenkinson, M. W. Woolrich, C. F. Beckmann, T. E. J. Behrens, H. Johansen-Berg, P. R. Bannister, M. De Luca, I. Drobnjak, D. E. Flitney, R. K. Niazy, J. Saunders, J. Vickers, Y. Zhang, N. De Stefano, J. M. Brady, P. M. Matthews. Advances in functional and structural mr image analysis and implementation as fsl. NeuroImage. Vol. 23, pg. S208–S219, 2004, https://doi.org/10.1016/j.neuroimage.2004.07.051. [↩]
- E. T. Rolls, C.-C. Huang, C.-P. Lin, J. Feng, M. Joliot. Automated anatomical labelling atlas 3. NeuroImage. Vol. 206, pg. 116189, 2020, https://doi.org/10.1016/j.neuroimage.2019.116189. [↩]
- C.-G. Yan, X.-D. Wang, X.-N. Zuo, Y.-F. Zang. DPABI: data processing & analysis for (resting-state) brain imaging. Neuroinformatics. Vol. 14, pg. 339–351, 2016, https://doi.org/10.1007/s12021-016-9299-4. [↩]
- S. Whitfield-Gabrieli, A. Nieto-Castanon. Conn: a functional connectivity toolbox for correlated and anticorrelated brain networks. Brain Connectivity. Vol. 2, pg. 125–141, 2012, https://doi.org/10.1089/brain.2012.0073. [↩]
- A. Zalesky, A. Fornito, E. T. Bullmore. Network-based statistic: identifying differences in brain networks. Neuroimage. Vol. 53, pg. 1197–1207, 2010, https://doi.org/10.1016/j.neuroimage.2010.06.041. [↩]
- JASP Team. JASP (version 0.95.1). 2025. [↩]
- J. L. Lanciego, N. Luquin, J. A. Obeso. Functional neuroanatomy of the basal ganglia. Cold Spring Harbor Perspectives in Medicine. Vol. 2, pg. a009621–a009621, 2012, [↩]
- W. Schultz. Reward functions of the basal ganglia. Journal of neural transmission (Vienna, Austria: 1996), Vol. 123, pg. 679–693, 2016, https://doi.org/10.1007/s00702-016-1510-0. [↩]
- Y. T. Cho, S. Fromm, A. E. Guyer, A. Detloff, D. S. Pine, J. L. Fudge, M. Ernst. Nucleus accumbens, thalamus and insula connectivity during incentive anticipation in typical adults and adolescents. NeuroImage. Vol. 66, pg. 508–521, 2013, https://doi.org/10.1016/j.neuroimage.2012.10.013. [↩]
- D. Zeithamova, B. D. Gelman, L. Frank, A. R. Preston. Abstract representation of prospective reward in the hippocampus. The Journal of Neuroscience. Vol. 38, pg. 10093–10101, 2018, https://doi.org/10.1523/jneurosci.0719-18.2018. [↩]
- T. H. Cheung & R. N. Cardinal. Hippocampal lesions facilitate instrumental learning with delayed reinforcement but induce impulsive choice in rats. BMC neuroscience. Vol. 6, pg. 36, 2005, https://doi.org/10.1186/1471-2202-6-36. [↩]
- B. W. Balleine, M. R. Delgado, O. Hikosaka. The role of the dorsal striatum in reward and decision-making. Journal of Neuroscience. Vol. 27, pg. 8161–8165, 2007, https://doi.org/10.1523/jneurosci.1554-07.2007. [↩]
- N. F. Parker, C. M. Cameron, J. P. Taliaferro, J. Lee, J. Y. Choi, T. J. Davidson, N. D. Daw, I. B. Witten. Reward and choice encoding in terminals of midbrain dopamine neurons depends on striatal target. Nature Neuroscience. Vol. 19, pg. 845–854, 2016, https://doi.org/10.1038/nn.4287. X. [↩]
- Gao, M. Zhang, Z. Yang, M. Wen, H. Huang, R. Zheng, W. Wang, Y. Wei, J. Cheng, S. Han, Y. Zhang. Structural and functional brain abnormalities in internet gaming disorder and attention-deficit/hyperactivity disorder: a comparative meta-analysis. Frontiers in Psychiatry. Vol. 12, 2021, https://doi.org/10.3389/fpsyt.2021.679437. [↩]
- H. Dong, M. Wang, J. Zhang, Y. Hu, M. N. Potenza, G. Dong. Reduced frontostriatal functional connectivity and associations with severity of internet gaming disorder. Addiction Biology. Vol. 26, 2020, https://doi.org/10.1111/adb.12985. [↩]
- K. Basar, T. Sesia, H. Groenewegen, H. W. M. Steinbusch, V. Visser-Vandewalle, Y. Temel. Nucleus accumbens and impulsivity. Progress in Neurobiology. Vol. 92, pg. 533–557, 2010, https://doi.org/10.1016/j.pneurobio.2010.08.007. [↩]
- L. Zhu, Y. Zhu, S. Li, Y. Jiang, M. Xian, Y. Wang, D. Yang, J. Zhao, L. Mu, W. Wang. Association of internet gaming disorder with impulsivity: role of risk preferences. BMC Psychiatry. Vol. 23, 2023, [↩]
- H. Ryu, J.-Y. Lee, A. Choi, S. Park, D.-J. Kim, J.-S. Choi. The relationship between impulsivity and internet gaming disorder in young adults: mediating effects of interpersonal relationships and depression. International Journal of Environmental Research and Public Health. Vol. 15, pg. 458, 2018, https://doi.org/10.3390/ijerph15030458. [↩]
- X. Xu, H. Ni, H. Wang, T. Wang, C. Liu, X. Song, G.-H. Dong. Functional connectivity-related changes underlying mindfulness meditation for internet gaming disorder: a randomized clinical trial. Npj Mental Health Research. Vol. 4, 2025, https://doi.org/10.1038/s44184-025-00154-6. [↩]
- C. Cai, K. Yuan, J. Yin, D. Feng, Y. Bi, Y. Li, D. Yu, C. Jin, W. Qin, J. Tian. Striatum morphometry is associated with cognitive control deficits and symptom severity in internet gaming disorder. Brain Imaging and Behavior. Vol. 10, pg. 12–20, 2015, https://doi.org/10.1007/s11682-015-9358-8. [↩]
- Y. Sun, H. Ying, R. M. Seetohul, W. Xuemei, Z. Ya, L. Qian, X. Guoqing, S. Ye. Brain fmri study of crave induced by cue pictures in online game addicts (male adolescents). Behavioural Brain Research. Vol. 233, pg. 563–576, 2012, https://doi.org/10.1016/j.bbr.2012.05.005. [↩]
- G.-C. Liu, J.-Y. Yen, C.-Y. Chen, C.-F. Yen, C.-S. Chen, W.-C. Lin, C.-H. Ko. Brain activation for response inhibition under gaming cue distraction in internet gaming disorder. The Kaohsiung Journal of Medical Sciences. Vol. 30, pg. 43–51, 2014, https://doi.org/10.1016/j.kjms.2013.08.005. [↩]
- A. Ciricugno, C. Ferrari, L. Battelli, Z. Cattaneo. A chronometric study of the posterior cerebellum’s function in emotional processing. Current Biology. Vol. 34, pg. 1844-1852.e3, 2009, https://doi.org/10.1016/j.cub.2024.03.013. [↩]
- C. Stoodley, J. Schmahmann. Functional topography in the human cerebellum: a meta-analysis of neuroimaging studies. NeuroImage. Vol. 44, pg. 489–501, 2009, https://doi.org/10.1016/j.neuroimage.2008.08.039. [↩]
- H. Wang, C. Jin, K. Yuan, T. M. Shakir, C. Mao, X. Niu, C. Niu, L. Guo, M. Zhang. The alteration of gray matter volume and cognitive control in adolescents with internet gaming disorder. Frontiers in Behavioral Neuroscience. Vol. 9, 2015, https://doi.org/10.3389/fnbeh.2015.00064. [↩]
- A.C. Bostan, P.L Strick. The Cerebellum and Basal Ganglia are Interconnected. Neuropsychol Rev. Vol. 20, pg. 261–270, 2010, https://doi.org/10.1007/s11065-010-9143-9. [↩]
- X. Li, H. Su, N. Zhong, T. Chen, J. Du, K. Xiao, D. Xu, W. Song, H. Jiang, M. Zhao. Aberrant resting-state cerebellar-cerebral functional connectivity in methamphetamine-dependent individuals after six months abstinence. Frontiers in Psychiatry. Vol. 11, 2020, https://doi.org/10.3389/fpsyt.2020.00191. [↩]
- M. Abdallah, N. M. Zahr, M. Saranathan, N. Honnorat, N. Farrugia, A. Pfefferbaum, E. V. Sullivan, S. Chanraud. Altered cerebro-cerebellar dynamic functional connectivity in alcohol use disorder: a resting-state fmri study. The Cerebellum. Vol. 20, pg. 823–835, 2021, [↩]
- L. C. Rice, M. T. Langan, D. T. Cheng, Y. Sheu, J. Peterburs, J. Hua, Q. Qin, J. J. Rilee, M. L. Faulkner, J. R. Mathena, C. A. Munro, G. S. Wand, M. E. McCaul, J. E. Desmond. Disrupted executive cerebro‐cerebellar functional connectivity in alcohol use disorder. Alcohol Clinical and Experimental Research. Vol. 48, pg. 33–47, 2023, https://doi.org/10.1111/acer.15219. [↩]
- N. Amokrane, C.-Y. R. Lin, N. A. Desai, S.-H. Kuo. The impact of compulsivity and impulsivity in cerebellar ataxia: a case series. Tremor and Other Hyperkinetic Movements. Vol. 10, pg. 43, 2020, https://doi.org/10.5334/tohm.550. [↩]
- J. He, H. Zhao, X. Lei, J. Qiu, T. Feng, H. Chen, O. Turel, A. Bechara, Q. He. How impulsivity and sensation seeking drive igd tendency in healthy young adults: a resting-state mri study. Journal of Behavioral Addictions. 2025, https://doi.org/10.1556/2006.2025.00063. [↩]
- J.-E. Jeong, C. Park, M. Kim, H. Cho, Arom Pyeon, S. Jung, D. Jung, J.-Y. Kim, J. Choi, J.-W. Chun, K.-J. Ahn, D.-J. Kim. Effects of bilateral tdcs over dlpfc on response inhibition, craving, and brain functional connectivity in internet gaming disorder: a randomized, double-blind, sham-controlled trial with fmri. Journal of Behavioral Addictions. Vol. 13, pg. 610–621, 2024, https://doi.org/10.1556/2006.2024.00017. [↩]
- A.-C. Schaub, M. Kirschner, N. Schweinfurth, L. Mählmann, C. Kettelhack, E. E. Engeli, J. P. K. Doll, S. Borgwardt, U. E. Lang, S. Kaiser, M. Walter, M. Herdener, J. Wrege, A. Schmidt. Neural mapping of anhedonia across psychiatric diagnoses: a transdiagnostic neuroimaging analysis. NeuroImage: Clinical. Vol. 32, pg. 102825, 2021 https://doi.org/10.1016/j.nicl.2021.102825. [↩]
- S. Muflih, S. I. Al-Azzam, K. H. Alzoubi, R. Karasneh, S. Hawamdeh, W. M. Sweileh. A bibliometric analysis of global trends in internet addiction publications from 1996 to 2022. Informatics in Medicine Unlocked. Vol. 47, pg. 101484, 2024 https://doi.org/10.1016/j.imu.2024.101484 [↩]



{kind=link}