
Sreenivas Eadara
Legend High School
Peer Reviewer: Manasi Soman
Pro Reviewer: Dr. Levente Pap
Abstract
There is a tremendous need for new agricultural developments to solve the iron deficiency anemia crisis. One such solution is examined here in the RedRice project; genetically modified rice can supplement heme iron as well as Vitamin B12 to treat populations worldwide. The project examined the procedures necessary to produce a genetically modified Oryza sativa (rice) plant using Agrobacterium tumefaciens genetic modification techniques. The genes transferred to the rice plant would include a leghemoglobin (heme iron protein) gene from Glycine max (soybean), as well as all genes of the Vitamin B12 pathway from Pseudomonas denitrificans. The Agrobacterium plasmid to be used was assessed.
Introduction
About 2 billion people worldwide are affected by anemia, a lack of red blood cells and reduced capacity to deliver oxygen to body organs (“Iron Deficiency Anaemia,” 2018). This is particularly common among poorer, undereducated individuals who know less about required nutrient intake or lack the funds necessary to purchase nutrient-rich foods. Anemia reduces the capacity of individuals to perform tasks while also increasing death rates in forms such as maternal hemorrhage. Those who suffer include individuals from Asian and African nations and developing nations where infectious diseases — which further complicate anemia — and poverty are common (“Iron Deficiency Anaemia,” 2018). Anemia is often caused by Iron or Vitamin B12 deficiencies.
Most anemia cases are due to Iron deficiency (“Iron Deficiency Anaemia,” 2018). Rice is the most common grain worldwide and is, therefore, the best vehicle for supplementation via genetic modification. Leghemoglobin, a form of hemoglobin like that in meats, is found in soybeans (Brisson and Verma, 1982). This modified rice, red in color, can be distributed in areas-in-need (the red color is introduced by the iron compounds, giving the rice a color like that of blood).
Vegetarians and the poor are particularly at-risk for anemia because they do not consume foods such as meats that are rich in iron and B12. RedRice gives such individuals opportunities to consume adequate amounts of iron (Watanabe, 2014).
Genetic Modification Techniques
Transformation, or the introduction of foreign DNA into a species’ genome, is the method by which these modifications are made. This transformation can be accomplished by several means. Agrobacterium tumefaciens, a bacterium that is usually known to cause crown gall disease on many plant species, can transfer a portion of its genome to plant cells, causing the development of crown gall disease symptoms within the plant (“Techniques for Genetically Modifying Plants,” n.d.). Different disarmed strains of Agrobacterium can transform target plant cells with portions of the soybean genome, forming genetically modified (GM) rice.
Among the methods considered for the transformation of rice were transformation by Agrobacterium, electroporation, and microprojectile bombardment (“Techniques for Genetically Modifying Plants,” n.d.). Unfortunately, Agrobacterium does not readily interact with rice, creating an obstacle to its use in this scenario. The electroporation process is inefficient and “requires laborious protocols” (Rivera et al., 2014).
Microprojectile bombardment, although effective, is quite expensive, and the bombardment process may damage cell DNA or introduce multiple copies of the transgenic DNA (Rivera et al., 2014). Therefore, it was most logical to proceed with the Agrobacterium tumefaciens transformation and to follow the best practices to increase the effectiveness of the method.
Naturally occurring plasmids within bacteria can be replicated independently from the bacterial chromosome, can carry specific genes that may confer specific traits to the bacteria, and can be communicated between bacteria themselves. Agrobacterium infects its hosts by the introduction of a Ti plasmid into the chromosomal DNA of host cells (“Techniques for Genetically Modifying Plants,” n.d.). Ti plasmids will first be isolated from Agrobacterium (J. Carlson, A. Stevens, J. Teichman, S. Fordham, personal communication, April 2, 2018). Afterward, restriction enzymes cut the plasmid DNA at certain loci (J. Carlson, A. Stevens, J. Teichman, S. Fordham, personal communication, April 2, 2018). After adding DNA with the same sticky ends produced by cutting other DNA with the same restriction enzymes, and “gluing” these fragments with DNA ligase, plasmids with foreign DNA, recombinant plasmids, are produced (J. Carlson, A. Stevens, J. Teichman, S. Fordham, personal communication, April 2, 2018). The plasmids can then be reintroduced to the Agrobacterium, and these bacteria can be introduced into a medium with rice cell protoplasts to transfer the recombinant plasmids to the plant cells (J. Carlson, A. Stevens, J. Teichman, S. Fordham, personal communication, April 2, 2018).
Supplemented Molecules
Leghemoglobin, an iron-containing oxygen binding compound found in Glycine Max (soybean) is easily processed by the body to produce hemoglobin. Hemoglobin is an essential oxygen-binding compound found in red blood cells essential for oxygen transport in the bloodstream; leghemoglobin is used in soybeans to prevent oxygen from interfering with enzymes and pathways required to “fix” nitrogen in the root nodules, where symbiotic bacteria cooperate with plant cells (Ott). B12, however, is a coenzyme, or a compound that assists with bodily chemical reactions. An essential amino acid, methionine, is synthesized by the MetH enzyme. This enzyme requires the presence of B12 for proper function, and without B12, this enzyme cannot function (Croft et al., 2006). B12 is produced naturally by Pseudomonas denitrificans in an aerobic manner (Caspi et al., 2013).
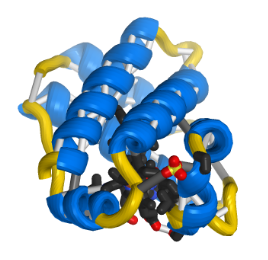
Figure 1. Leghemoglobin A.
This graphic was automatically generated through the NIH 3D print service (Coakley et al., 2014). It depicts a ribbon model for the 3D structure of the leghemoglobin a protein as determined by an X-ray diffraction.
This graphic was automatically generated through the NIH 3D print service (Coakley et al., 2014). It depicts a ribbon model for the 3D structure of the leghemoglobin a protein as determined by an X-ray diffraction.
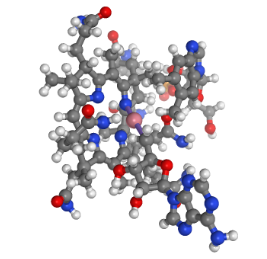
Figure 2. Vitamin B12.
This graphic was generated through the NIH 3D Print service using files obtained from ChemSpider (Coakley et al., 2014; “Cobamamide,” n.d.). It depicts a ball-and-stick model for the structure of Vitamin B12.
The product carries a distinct advantage because it produces heme iron, which is more bioavailable than the ferritin iron found in other plants. Ferritin iron absorption proves troublesome; it must be hydrolyzed and then made soluble, requiring the presence of hydrochloric acid in the stomach and Vitamin C in food (Bejjani et al., 2007). These constraints pose significant hurdles for its use. However, heme iron can be absorbed into the bloodstream in two ways, the first involving direct absorption. The second involves the conversion of heme iron into ferrous iron, which then must also convert to ferric iron, so it can bind to apotransferrin (West and Oates, 2008). Ferritin, however, must always follow a single pathway.
Materials + Methods
The processes and intuitions behind the genetic modification process necessary to produce this pioneering crop are explained below.
Transformation
Many Ti plasmids function via a binary vector system, which allows two plasmids to be used in the transformation process. One plasmid will store several of the sequences necessary for transformation, and the other will store other necessary frequencies as well as the DNA segments to be transferred to the infected plant cells (Gordon and Christie, 2014). PGreen0029 and pSoup form one such binary vector system that is both efficient and popular (Gordon and Christie, 2014). Common issues with vector systems include failures to insert sequences into plasmids, failure of bacteria to take up these sequences, and failures of the Agrobacterium to transform plant cells (Cirpus et al., 2005).
PGreen0029 facilitates verification that sequences were inserted by means of blue-white selection; restriction enzymes cut at a Multiple Cloning Site in the middle of a lac gene responsible for the processing of lactose (Hellens et al., 2007). If the sequence was inserted before ligation, the lac gene becomes unusable; if not, the lac gene continues to function. When cultured on a plate filled with kanamycin, an antibiotic, and X-gal, which forms a blue product when processed by bacteria with functional lac genes, E. coli (DH5 alpha competent strain) that did not pick up a pGreen0029 plasmid (which has a kanamycin resistance gene) will die, while E. coli that picked up a pGreen0029 plasmid will survive (Reece et al.). Those that survive will form white or blue colonies if they picked up a plasmid that did or did not integrate a new segment of the desired DNA after the restriction cut (Reece et al.). This process permits verification that sequences were inserted into the DH5? strain (J. Carlson, A. Stevens, J. Teichman, S. Fordham, personal communication, April 2, 2018). These bacteria can then replicate pGreen0029 (pGreen0029 is a high copy plasmid) and then a miniprep process can be used to extract the plasmids from the bacteria (J. Carlson, A. Stevens, J. Teichman, S. Fordham, personal communication, April 2, 2018). These plasmids, when introduced to Agrobacterium along with pSoup, can then be used for transformation.
When rice protoplasts are transformed and the plants regenerate, a selectable marker (antibiotic resistance) that was conferred to the plant via pGreen0029 and pSoup (kanamycin) will distinguish transformed plants from most other plants since they should not survive the antibiotic (J. Carlson, A. Stevens, J. Teichman, S. Fordham, personal communication, April 2, 2018). Sometimes, during the infection process, T-DNA towards the RB becomes significantly eroded (Lee and Gelvin, 2008). PGreen0029 combats this effect with 405bp of the lac gene and other sequences that put distance between the MCS and the RB (Benchling, 2018). Gibberellins, a class of plant hormone, can be used to speed up plant growth for laboratory purposes and allow verification that leghemoglobin and B12 are produced (J. Carlson, A. Stevens, J. Teichman, S. Fordham, personal communication, April 2, 2018). PSoup can be purchased within the popular Agrobacterium strain GV3101 for simplicity. GV3101 has chromosomal rifampin resistance, but this will not impede any selective processes since kanamycin, not rifampin, is used as a selectable marker (Lee and Gelvin, 2008).
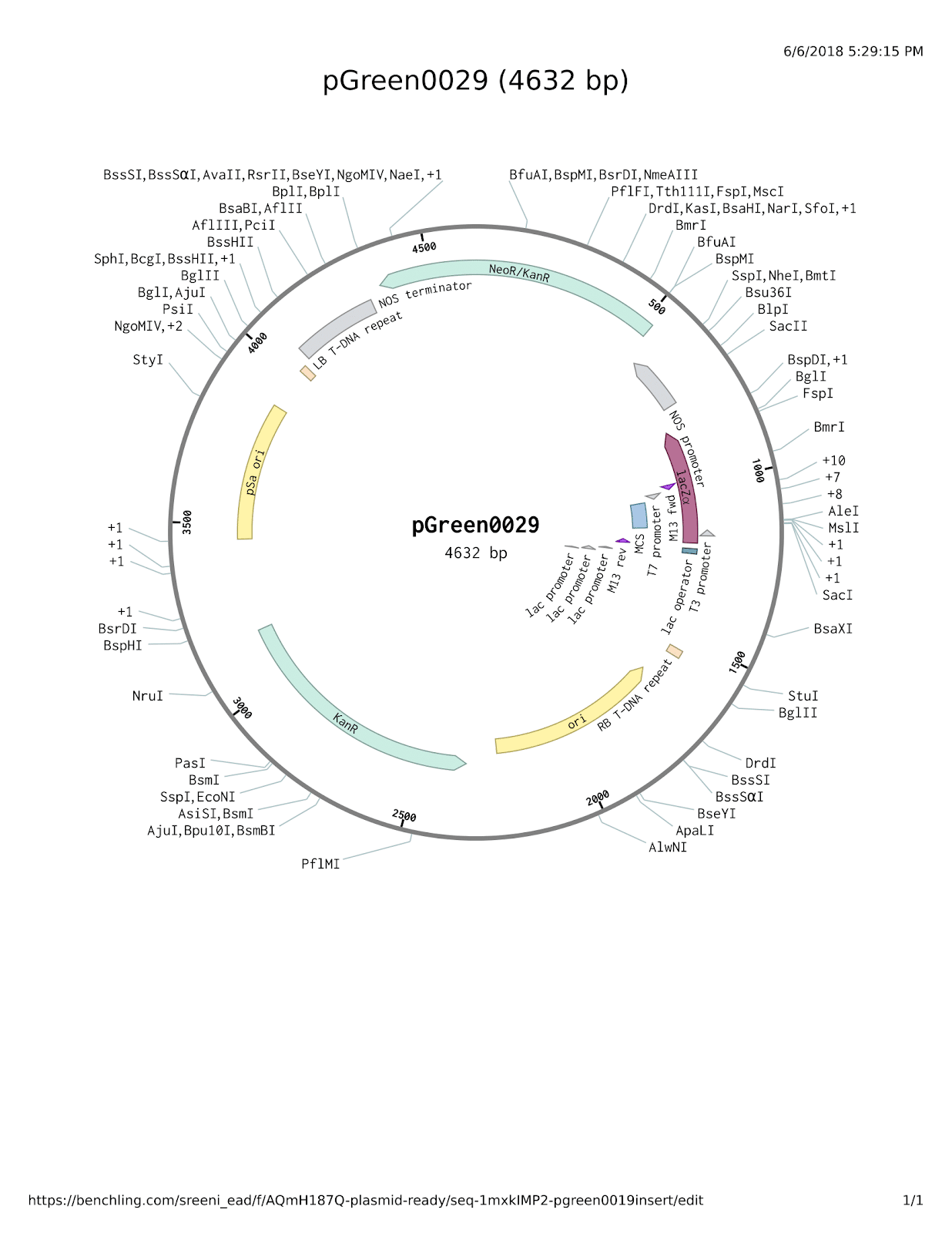
Figure 1. The pGreen0029 plasmid.
Plasmid structure was obtained from SnapGene and then modified as necessary in Benchling (SnapGene software (from GSL Biotech; available at snapgene.com), 2018; Benchling, 2018). Modifications can be seen in figures 5 and 6. Displayed below is the plasmid without any modification.
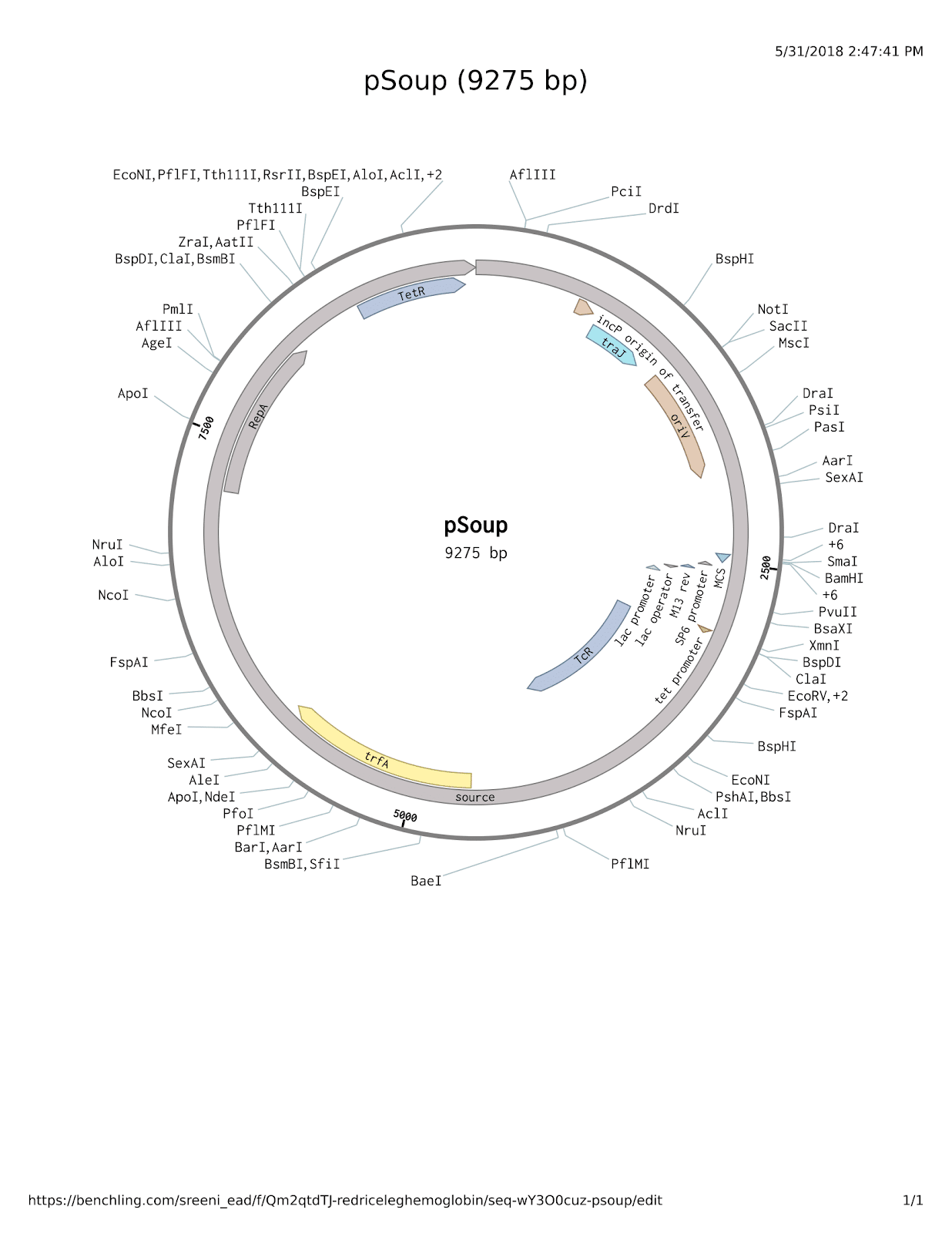
Figure 2. The pSoup plasmid.
Plasmid structure was obtained from SnapGene and the image was generated in Benchling (SnapGene software (from GSL Biotech; available at snapgene.com), 2018; Benchling, 2018). Displayed below is the pSoup plasmid without any modification.
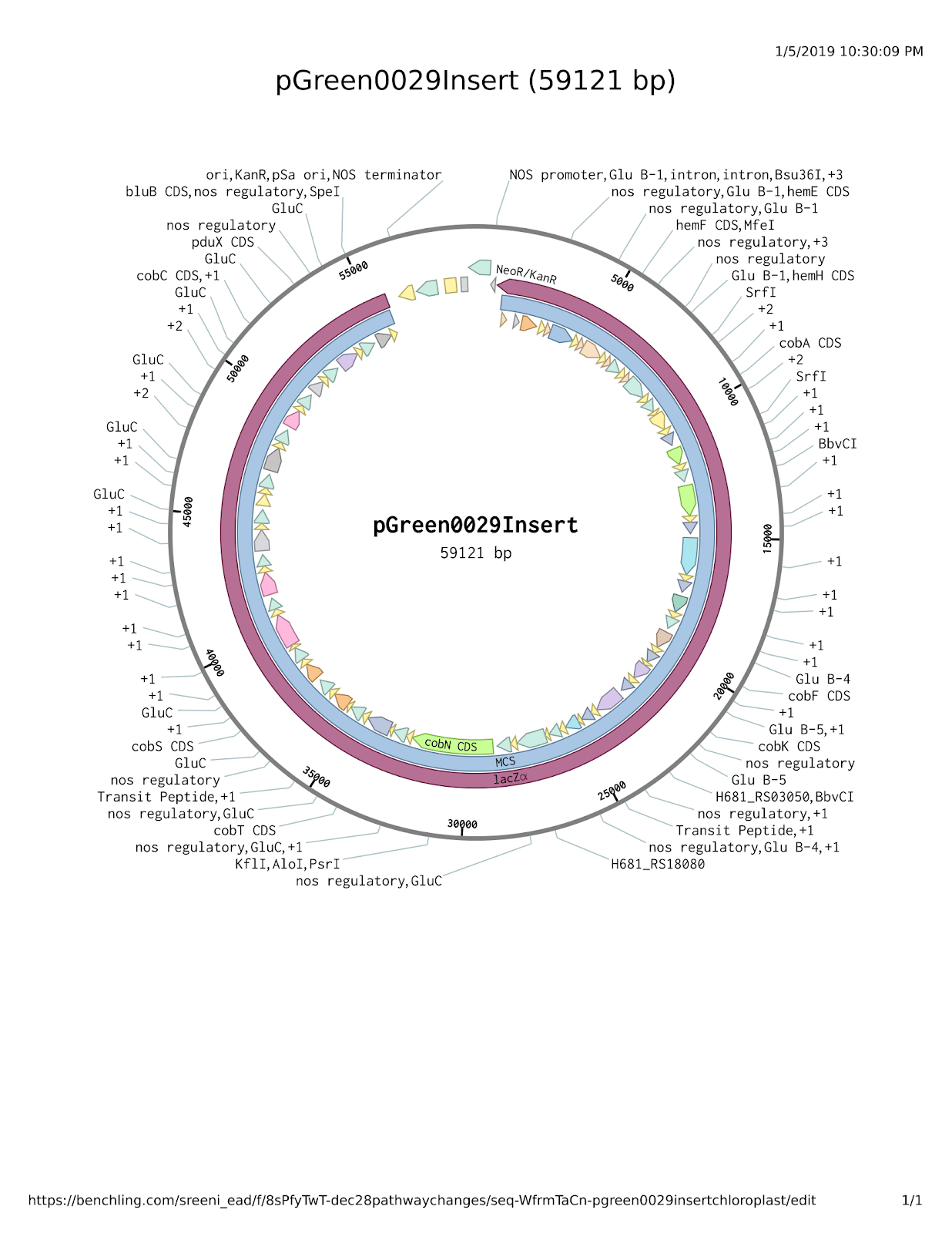
Figure 3. The pGreen0029 Plasmid with Complete Insert.
This graphic was generated in Benchling (2018). Nucleotide sequences were obtained from NCBI and were then added to the appropriate location within the insert (O’Leary et al., 2016). Each gene displayed was fitted with a promoter and terminator for seed-specific expression in the rice.
Genetic Composition
Several genes related to the production of leghemoglobin in soybeans and other genes related to the production of Vitamin B12 were located. However, since the expression of these genes involves the transferring of sequences, usually expressed in prokaryotes to plants, several steps must be taken to ensure that the sequences are effective in their new location within the modified rice genome. These genes must be equipped with the necessary regulatory sequences to ensure that they can properly interact with eukaryotic gene regulation mechanisms, including necessary polymerase recognition sequences on both the 5’ and 3’ ends (Campbell and Reece, 2008, p. 359). Southern hybridization can then confirm that the sequences were accurately inserted (Fraley et al., 1983). These sequences can become effective if they expressed in the rice endosperm for human consumption via an endosperm-specific promoter. The Glu promoter is a promoter specific to the endosperm of rice, which will allow the Vitamin B12 and leghemoglobin production to be present in consumed rice (Ye et al., 2000). The endosperm is the content of seeds which provides for the development of the seeds once they are planted in fertile soil and constitutes most of the rice grain (Campbell and Reece, 2008, p. 807).
However, since longer insert sequences reduce the copy number of the plasmid, it is imperative that the length of our insert is as small as possible while maintaining effectiveness (“Plasmids 101: A Desktop Resource (3rd Edition)”, 2017). Wu et al. studied the effects of reducing promoter length of Glu B-1 on the activity in seed endosperm (1998). It was determined that the GCN4 motif was most necessary for the expression in the endosperm, while the ACAA motif and preceding sequences were only partially impactful on endosperm-specific expression. It was decided that a 2,292 bp promoter would unnecessarily impede the copy number of the pGreen0029 plasmid once the insert was introduced and instead a 245 bp segment of the promoter would be used, containing the GCN4 motif as well as two ACAA motifs.
It was noted that the repetition of several promoter or terminator sequences could lead to the instability or recombination of T-DNA (Cirpus et al., 2005). To prevent this result, it was decided that the promoter sequences used must be of differing sequences. However, there was another risk: if the sequences of the promoters used to regulate B12 pathway were not similar enough, the enzymes used to catalyze the aerobic production of B12 would not be produced in the same conditions. A compromise was reached by the introduction of several different promoters in the Glu gene family, including Glu B-4, B-5, and Glu C (Xu et al., 2016). These endosperm-specific promoter sequences are quite active and are expressed throughout the endosperm, while other genes in the Glu A and Glu B families are expressed at different times in different sections of endosperm (Xu et al., 2016). These sequences are similarly reduced in size to reduce insert size. To preserve the activity of the promoter, the B-4, B-5, and C promoters are shortened to 500bp and 600bp respectively. The nos terminator will still, however, be used for all genes.
A nos terminator sequence is often necessary to cause the rice cells’ translation machinery to cease “reading” the DNA transcript (“Plasmids 101: A Desktop Resource (3rd Edition)”, 2017). To ensure that the gene is properly transcribed, a decision was made to replace the terminator sequence common to the LBA gene with the nos terminator from A. tumefaciens. Used in the Golden Rice Project and many others as a sequence for the termination of transgene transcripts, it has proven its merit (Ye et al, 2000).
The Lb gene family is a group of closely related leghemoglobin genes. The LBA, LBC1, LBC2, and LBC3 genes are functional and responsible for leghemoglobin production (Brisson and Verma, 1982). LBC3 transformation and later production of leghemoglobin in other species have proven successful (Stougaard, 1987). The production of Vitamin B12 in rice can be achieved through similar transformation methods with the CobA, CobI, CobG CobJ, CobM, CobF, CobK, CobL, CobH, CobB, CobN, CobS, CobT, CobO, CobQ, CobC, CobD, and CobP, CobU, CobV, PhpB, PduX, and BluB genes (Caspi et al., 2013). These genes will produce adenosylcobalamin, an active form of Vitamin B12. Adenosylcobalamin need only bind to an intrinsic factor to become absorbed through ileal receptors and into the bloodstream.
It is important to note, however, that leghemoglobin is a holoprotein that consists of both an apoprotein and a heme moiety (Santana et al., 1998). Doubts exist surrounding the true nature of heme moiety synthesis within the root nodules of Glycine Max, but as suggested by Mark O’Brian, that process involves protoporphyrin IX synthesis (O’Brian, 1996). E. coli contains a heme synthesis pathway containing this key intermediate through which it produced heme b aerobically. The genes necessary to produce heme include HemE, HemF, HemG, and HemH (Caspi et al., 2013). These genes were placed in the insert.
Uroporphyrinogen III is a molecule of interest as it is an intermediate in heme, chlorophyll, and B12 production (Dailey et al., 2017). Since Uroporphyrinogen III is produced in the stroma of the chloroplast during chlorophyll synthesis, it may indeed be possible to repurpose some Uroporphyrinogen III for Vitamin B12 and heme production within the stroma of the chloroplast (Chlorophyll A Synthesis). This is achieved by attaching a chloroplast transit peptide to each gene required for heme synthesis or Vitamin B12 synthesis (Ye et al., 2000). The transit peptide to be used is the pea RuBisCO small subunit transit peptide (Lübeck et al., 1997). This will allow the enzymes intended for the B12 and heme pathways to enter the stroma of the chloroplast (Lübeck et al., 1997). If this occurs, there is no longer a need to include genes necessary for Uroporphyrinogen III in the insert of pGreen0029. This can reduce the insert size.
As rice seeds finish development, chlorophyll degrades because of the activity of two reductases, a magnesium dechelatelase, and other enzymes (Smolikova et al., 1993). As chlorophyll degrades, heme and B12 should be unaffected, leaving their distinct red color. This occurs because the Mg2+ dechelatelase is specific to Mg2+ and the pheophytinase enzyme is extremely specific to pheophytin (Smolikova et al., 1993). Without the action of these enzymes and others, the splitting of the porphyrin ring and further degradation cannot occur (Smolikova et al., 1993).
Results + Discussion
If all goes successfully, a transgenic plant capable of leghemoglobin and Vitamin B12 production should be produced. The plant will produce heme via the E. coli heme pathway and Vitamin B12 via the P. aeruginosa pathway. It will produce the leghemoglobin apoprotein as directed by a G. max gene. The production of the leghemoglobin apoprotein could be verified via Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis, or SDS-PAGE (Team Oxford, 2014). This process isolates proteins from a sample of homogenized rice seed endosperm and runs denatured proteins through a gel to isolate them by size. Since the size of the leghemoglobin protein is known (15374 Da), imaging could reveal the concentration of the leghemoglobin apoprotein relative to the concentrations of other endosperm proteins (Benchling, 2018). The production of the heme moiety and Vitamin B12 would have to be verified via High Performance Liquid Chromatography (HPLC) to determine the relative concentration in absorbance of a specified wavelength. HPLC analysis will produce a chart with separate “peaks,” each indicating the absorbance of a compound. By comparing the peaks shown in HPLC of the endosperm with HPLC of pure Vitamin B12 and pure heme, we can determine which peaks within the endosperm correspond to which molecules and analyze their absorbance (“The Theory of HPLC, Quantitative and Qualitative HPLC,” n.d.). If the concentrations of leghemoglobin and Vitamin B12 are insufficient for dietary purposes, modification of the approach (which would most likely include modification of the promoter sequences to amplify production) would be necessary.
Potential Concerns & Areas for Improvement
One of the greatest concerns with the ATMT is the insert size and recombination risk. Since so many genes are involved in the B12 and heme pathways, a need to express these genes in the same locations and a need to prevent recombination had to be balanced, since promoter sequences were repeated. A tentative solution included the distribution of three different promoter sequences to drive the endosperm-specific production of the genes as described above. The extremely large insert size, however, could scarcely be avoided, and was only somewhat helped by shortening promoter sequences. The only available option would include the removal of the heme pathway from the insert and using another pathway native to the rice genome.
Rice does produce some of its own hemoglobins of the hb gene family of nonsymbiotic hemoglobins, including hb1, hb2, hb3, hb4, and hb5, which may indeed be functional as indicated by upstream promoters but regulated in a complex manner (Arredondo et al., 2018). Further investigation is needed to determine whether these rice hemoglobins can help treat anemia and whether the production of these rice hemoglobins can be accelerated within the seed (O’Brian, 1996). This would likely involve a CRISPR – Cas9 system for plant modification where the sequences necessary for heme and hemoglobin production would be identified and their promoters would be mutated such that their expression would be enhanced in rice seed endosperm (Visk, 2018). It is important to note that increasing the expression of these genes throughout the rice plant would be a detriment to the plant’s function; this pathway is usually evoked in the event stressors such as cold, drought, and salt stress within organs affected by such specific stressors (Arredondo et al., 2018).
Environmental activists have expressed significant concerns about the production and cultivation of GMOs like RedRice. They posit that the modifications found in GMOs have the potential to spread to native, non-modified species in a phenomenon termed transgene escape (Daniell, 2002). The potential for this is limited, however, since the pollen of rice is short-lived and the potential for crossing with wild-type rice is very low. Rice disperses via the seed, and if the seed is properly handled, potential environmental effects could be averted. Modifications to the reproductive system of the plant to reduce the potential for outcrossing and therefore transgene escape would not prevent this seed dispersal. The added pathways and genetic modifications should not modify the growth or reproductive characteristics of the rice plant. The new plant may even suffer a selective disadvantage since the presence of heme and B12 within the rice might make it more attractive to insects or other animals (Golden Rice Humanitarian Board).
Overall Impact
RedRice can save billions from anemia, and rice consumption is universal. In 2017 alone, it was estimated that 477.7 million metric tons of rice were consumed (“The importance of rice,” 2007). Of this, rice alone provides food to 21% of the total global human – per capita – energy consumption (“The importance of rice,” 2007). This worldwide resource could be tapped to allow the required daily intakes of iron to reach target populations. The advantages to worldwide health would be colossal. If individuals in high-risk areas consume as much as 7 milligrams of iron contained in RedRice, as well as the included Vitamin B12 daily, anemia could be avoided.
Target populations largely reside in the African continent, southern Asia, and middle South America (“Micronutrient Deficiencies,” n.d.). Rice is a staple grain in Asia and “the most important commodity in their daily lives” (“The importance of rice,” 2007). This makes it well suited to those populations. Brazil, which is at the heart of the anemic South American population, ranks the 14th in global rice consumption, making RedRice a useful tool in addressing anemia in that area as well.
Conclusion
RedRice can be a versatile tool in the treatment of iron and Vitamin B12 deficiencies and therefore nutrient deficiency anemia. The insert within the pGreen0029 plasmid contains all the genes specific to the leghemoglobin and B12 pathways, including the heme pathway necessary for leghemoglobin production. The expression of these genes was directed via highly active endosperm-specific promoters, and different promoters were selected to help prevent destabilization of the insert DNA as the sequences are repetitive (Xu et al.; Cirpus et al., 2005). Heme and Vitamin B12 production was directed towards the chloroplast, as the chlorophyll intermediate Uroporphyrinogen III can be utilized in those pathways (Chlorophyll A Synthesis). With the completed pGreen0029 plasmid and included insert, as well as the pSoup plasmid, Agrobacterium can create transgenic rice capable of Vitamin B12 and leghemoglobin production (Gordon and Christie, 2014; Cirpus et al., 2005).
RedRice, as a genetically modified organism, possesses little risk for transgene escape or environmental damages (Golden Rice Humanitarian Board). This, when considered alongside the prevalence of rice as a staple food of populations worldwide, makes RedRice incredibly well-suited to the treatment of anemia on a global scale (“The importance of rice,” 2007). Targets for RedRice would include the African continent, southern Asia, and middle South America (“Micronutrient Deficiencies,” n.d.).
Potential refinements may depend on further research into the production of heme in non-transgenic rice. By adapting natural heme production to leghemoglobin production within endosperm, the need for the E. coli heme pathway is eliminated and the insert size is reduced. This rice heme pathway, however, has yet to be completely characterized (Arredondo et al., 2018).
Nevertheless, the RedRice project has the potential to save billions of lives from iron and Vitamin B12-deficiency anemias via daily consumption.
Works Cited
- Arredondo-Peter, R., Moran, J. F., & Sarath, G. (2014, December 11). Rice (Oryza) hemoglobins. F 1000 research, 3(253). doi:10.12688/f1000research.5530.2
- Ayliffe, M. A., Agostino, A., Clarke, B. C., Furbank, R., Caemmerer, S. V., & Pryor, A. J. (2009, March). Suppression of the Barley uroporphyrinogen III synthase Gene by a Ds Activation Tagging Element Generates Developmental Photosensitivity. The Plant Cell Online, 21(3), 814-831. doi:10.1105/tpc.108.063685
- Becana Ausejo, M., Morán, J. F., Iturbe-Ormaetxe, I., Gogorcena Aoiz, Y., & Escuredo, P. R. (1995). Structure and function of leghemoglobins.
- Bejjani, S., Pullakhandam, R., Punjal, R., & Nair, K. (2007). Gastric digestion of pea ferritin and modulation of its iron bioavailability by ascorbic and phytic acids in caco-2 cells. World Journal of Gastroenterology, 13(14), 2083. doi:10.3748/wjg.v13.i14.2083
- Benchling [Biology Software]. (2018). Retrieved from https://benchling.com.
- Brisson, N., & Verma, D. P. (1982). Soybean leghemoglobin gene family: Normal, pseudo, and truncated genes. Proceedings of the National Academy of Sciences, 79(13), 4055-4059. doi:10.1073/pnas.79.13.4055
- Campbell, N. A., & Reece, J. B. (2008). Biology. San Francisco: Pearson Benjamin Cummings.
- Caspi, R., Altman, T., Billington, R., Dreher, K., Foerster, H., Fulcher, C. A., … Karp, P. D. (2013). The MetaCyc database of metabolic pathways and enzymes and the BioCyc collection of Pathway/Genome Databases. Nucleic Acids Research, 42(D1). doi:10.1093/nar/gkt1103
- Chlorophyll A Synthesis. (n.d.). Retrieved from https://sites.tufts.edu/photosyntheticanimals/biochemical-pathway/
- Choby, J. E., & Skaar, E. P. (2016, August 28). Heme Synthesis and Acquisition in Bacterial Pathogens. Journal of Molecular Biology, 428(17), 3408-3428. doi:10.1016/j.jmb.2016.03.018
- Cirpus, P., Bauer, J., Qui, X., Wu, G., & Datla, N. (2005). U.S. Patent No. 10590457. Washington, DC: U.S. Patent and Trademark Office.
- Claros M.G., Vincens P. (1996). Computational method to predict mitochondrially imported proteins and their targeting sequences. Eur. J. Biochem. 241, 770-786.
- Coakley, M.F., Hurt, D.E., Weber, N., Mtingwa, M., Fincher, E.C., Alekseyev, V., … Huyen, Y. (September 2014). The NIH 3D print exchange: A public resource for bioscientific and biomedical 3D prints. 3D Printing and Additive Manufacturing, 1(3), 137-140. doi: 10.1089/3dp.2014.1503.
- Cobamamide. (n.d.). Retrieved from http://www.chemspider.com/Chemical-Structure.16736117.html?rid=677d1369-320c-430b-a8f5-37c375dd834f&page_num=0
- Coleman, E. (2017, December 6). The recommended amount & percent of carbohydrates per day. Retrieved from https://healthyeating.sfgate.com/recommended-amount-percent-carbohydrates-per-day-7287.html
- Croft, M. T., Warren, M. J., & Smith, A. G. (2006). Algae need their vitamins. Eukaryotic Cell, 5(8), 1175-1183. doi:10.1128/ec.00097-06
- Dailey, H. A., Dailey, T. A., Gerdes, S., Jahn, D., Jahn, M., Obrian, M. R., & Warren, M. J. (2017). Prokaryotic heme biosynthesis: Multiple pathways to a common essential product. Microbiology and Molecular Biology Reviews, 81(1). doi:10.1128/mmbr.00048-16
- Daniell, H. (2002). Molecular strategies for gene containment in transgenic crops. Nature Biotechnology, 20(6), 581-586. doi:10.1038/nbt0602-581
- Dietary Supplement Fact Sheet: Iron. (2018, March 2). Retrieved from http://ods.od.nih.gov/factsheets/Iron-HealthProfessional/
- Eckardt, N. A. (2006). Cytoplasmic male sterility and fertility restoration. The Plant Cell Online, 18(3), 515-517. doi:10.1105/tpc.106.041830
- Fabregat, A., Jupe, S., Matthews, L., Sidiropoulos, K., Gillespie, M., Garapati, P., … D’Eustachio, P. (2017, November 14). The reactome pathway knowledgebase. Nucleic Acids Research, 46(D1). doi:10.1093/nar/gkx1132
- Fang, H., Kang, J., & Zhang, D. (2017). Microbial production of vitamin B12: A review and future perspectives. Microbial Cell Factories, 16(1). doi:10.1186/s12934-017-0631-y
- Fraley, R. T., Rogers, S. G., Horsch, R. B., Sanders, P. R., Flick, J. S., Adams, S. P., … Woo, S. C. (1983). Expression of bacterial genes in plant cells. Proceedings of the National Academy of Sciences, 80(15), 4803-4807. doi:10.1073/pnas.80.15.4803
- Golden Rice Humanitarian Board. Golden rice risk assessment. (http://www.goldenrice.org/Content2-How/how3a_biosafety.php)
- Gordon, J. E., & Christie, P. J. (2014). The Agrobacterium Ti plasmids. Microbiology Spectrum, 2(6). doi:10.1128/microbiolspec.plas-0010-2013
- Group, E. (2017, February 22). 4 must know facts about Adenosylcobalamin. Retrieved from http://www.globalhealingcenter.com/natural-health/adenosylcobalamin-4-facts-know/
- Hamilton, C. M. (1997). A binary-BAC system for plant transformation with high-molecular-weight DNA. Gene, 200(1-2), 107-116. doi:10.1016/s0378-1119(97)00388-0
- Hellens, R. P., Edwards, E. A., Leyland, N. R., Bean, S., & Mullineaux, P. M. PGreen: A versatile and flexible binary Ti vector for Agrobacterium-mediated plant transformation. (2000). Plant Molecular Biology, 42(6), 819-832. doi:10.1023/a:1006496308160
- Helliwell, K. E., Scaife, M. A., Sasso, S., Araujo, A. P., Purton, S., & Smith, A. G. (2014, March 13). Unraveling vitamin B12-responsive gene regulation in algae. Plant Physiology, 165(1), 388-397. doi:10.1104/pp.113.234369
- Helliwell, K., Lawrence, A., Holzer, A., Kudahl, U., Sasso, S., Kräutler, B., … Smith, A. (2016, April 25). Cyanobacteria and eukaryotic algae use different chemical variants of vitamin B12. Current Biology, 26(8), 999-1008. doi:10.1016/j.cub.2016.02.041
- Hörtensteiner, S. (2012, July 13). Update on the biochemistry of chlorophyll breakdown. Plant Molecular Biology, 82(6), 505-517. doi:10.1007/s11103-012-9940-z
- Hoy, J. A., & Hargrove, M. S. (2008, March). The structure and function of plant hemoglobins. Plant Physiology and Biochemistry, 46(3), 371-379. doi:10.1016/j.plaphy.2007.12.016
- Importance of Rice. (2007). Retrieved from http://www.knowledgebank.irri.org/ericeproduction/Importance_of_Rice.htm
- Iron Deficiency Anaemia. (n.d.). Retrieved February 13, 2018, from www.who.int/nutrition/topics/ida/en/
- Kaewsuksaeng, S. (2011). Chlorophyll degradation in horticultural crops. Walailak Journal of Science and Technology (WJST), 8(1), 9-19.
- Kyker, K. (n.d.). Protoplast production. Retrieved from http://ind.ntou.edu.tw/~b0232/proto2.htm
- Layer, G., Reichelt, J., Jahn, D., & Heinz, D. W. (2010, April 27). Structure and function of enzymes in heme biosynthesis. Protein Science, 19(6), 1137-1161. doi:10.1002/pro.405
- Lee, L., & Gelvin, S. B. (2008). T-DNA binary vectors and systems. Plant Physiology, 146(2), 325-332. doi:10.1104/pp.107.113001
- Lübeck, J., Heins, L., & Soll, J. (1997). A Nuclear-coded chloroplastic inner envelope membrane protein uses a soluble sorting intermediate upon import into the organelle. The Journal of Cell Biology, 137(6), 1279-1286. doi:10.1083/jcb.137.6.1279
- Nadler, K. D., & Avissar, Y. J. (1977, September). Heme synthesis in soybean root nodules. Plant Physiology, 60(3), 433-436. doi:10.1104/pp.60.3.433
- O’Leary, N.A., Wright, M.W., Brister, J.R., Ciufo, S., Haddad, D., McVeigh, R., … Pruitt, K.D. (2016, Jan 4). Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res, 44. doi:10.1093/nar/gkv1189
- O’Brian, M. R. (1996, May). Heme synthesis in the rhizobium-legume symbiosis: A palette for bacterial and eukaryotic pigments. Journal of Bacteriology, 178(9), 2471-2478. doi:10.1128/jb.178.9.2471-2478.1996
- O’Brian, M. R., Kirshbom, P. M., & Maier, R. J. (1987). Bacterial heme synthesis is required for expression of the leghemoglobin holoprotein but not the apoprotein in soybean root nodules. Proceedings of the National Academy of Sciences, 84(23), 8390-8393. doi:10.1073/pnas.84.23.8390
- Park, S., Yu, J., Park, J., Li, J., Yoo, S., Lee, N., … Paek, N. (2007, May). The senescence-induced staygreen protein regulates chlorophyll degradation. The Plant Cell Online, 19(5), 1649-1664. doi:10.1105/tpc.106.044891
- Plasmid Cloning by PCR. (n.d.). Retrieved from http://www.addgene.org/protocols/pcr-cloning/
- Plasmids 101: A Desktop Resource (3rd Edition). (2017). Retrieved from https://info.addgene.org/download-addgenes-ebook-plasmids-101-3rd-edition
- Pseudomonas Aeruginosa PAO1 Adenosylcobalamin Biosynthesis II (Late Cobalt Incorporation). (n.d.). Retrieved from http://www.pseudomonas.com:1555/PSEUDO/NEW-IMAGE?object=P381-PWY
- Rivera, A. L., Gómez-Lim, M., Fernández, F., & Loske, A. M. (2014). Genetic transformation of cells using physical methods. Journal of Genetic Syndromes & Gene Therapy, 05(04). doi:10.4172/2157-7412.1000237
- Rodionov, D. A., Vitreschak, A. G., Mironov, A. A., & Gelfand, M. S. (2003). Comparative genomics of the vitamin B12 metabolism and regulation in prokaryotes. Journal of Biological Chemistry, 278(42), 41148-41159. doi:10.1074/jbc.m305837200
- Sakuraba, Y., Kim, E., Han, S., Piao, W., An, G., Todaka, D., … Paek, N. (2017, July 11). Rice phytochrome-interacting factor-like1 (OsPIL1) is involved in the promotion of chlorophyll biosynthesis through feed-forward regulatory loops. Journal of Experimental Botany, 68(15), 4103-4114. doi:10.1093/jxb/erx231
- Santana, M. A., Pihakaski-Maunsbach, K., Sandal, N., Marcker, K. A., & Smith, A. G. (1998, April). Evidence that the plant host synthesizes the heme moiety of leghemoglobin in root nodules. Plant Physiology, 116(4), 1259-1269. doi:10.1104/pp.116.4.1259
- Shankar, S., & Hoyt, M. A. (2016). U.S. Patent No. WO2016183163A1. Washington, DC: U.S. Patent and Trademark Office.
- Smolikova, G., Dolgikh, E., Vikhnina, M., Frolov, A., & Medvedev, S. (2017, September 16). Genetic and hormonal regulation of chlorophyll degradation during maturation of seeds with green embryos. International Journal of Molecular Sciences, 18(9), 1993. doi:10.3390/ijms18091993
- Stougaard, J., Petersen, T. E., & Marcker, K. A. (1987). Expression of a complete soybean leghemoglobin gene in root nodules of transgenic Lotus corniculatus. Proceedings of the National Academy of Sciences, 84(16), 5754-5757. doi:10.1073/pnas.84.16.5754
- Team Oxford. (2014). SDS-PAGE. IGEM Foundation. Retrieved from www.2014.igem.org/wiki/images/d/d1/SDS-PAGE.pdf
- Techniques for genetically modifying plants. (n.d.). Retrieved from http://www.genewatch.org/sub-532324
- The theory of HPLC, Quantitative and qualitative HPLC. (n.d.) Crawford Scientific. Retrieved from https://www.chromacademy.com/lms/sco9/Theory_Of_HPLC_Quantitative_and_Qualitative_HPLC.pdf
- Treatment with high dose vitamin B12 been shown to be safe for more than 50 years. (2009, April 15). Retrieved from https://stichtingb12tekort.nl/wetenschap/stichting-b12-tekort-artikelen/english/treatment-with-high-dose-vitamin-b12-been-shown-to-be-safe-for-more-than-50-years/
- UniProt: The universal protein knowledgebase. (2016). Nucleic Acids Research, 45(D1). doi:10.1093/nar/gkw1099
- Verma, D. P., & Bal, A. K. (1976, November 01). Intracellular site of synthesis and localization of leghemoglobin in root nodules. Proceedings of the National Academy of Sciences, 73(11), 3843-3847. doi:10.1073/pnas.73.11.3843
- Virmani, S. S. (2003). Two-line hybrid rice breeding manual. IRRI.
- Visk, D. (2018, October 31). CRISPR Applications in Plants. Retrieved from https://www.genengnews.com/insights/crispr-applications-in-plants/
- Watanabe, F., Yabuta, Y., Bito, T., & Teng, F. (2014). Vitamin B12-containing plant food sources for vegetarians. Nutrients, 6(5), 1861-1873. doi:10.3390/nu6051861
- West, A. R., & Oates, P. S. (2008). Mechanisms of heme iron absorption: Current questions and controversies. World Journal of Gastroenterology, 14(26), 4101. doi:10.3748/wjg.14.4101
- Wettstein, D. V. (1995, July). Chlorophyll Biosynthesis. The Plant Cell Online, 7(7), 1039-1057. doi:10.1105/tpc.7.7.1039
- Woodson, J., Perez-Ruiz, J., & Chory, J. (2011, May 12). Heme synthesis by plastid Ferrochelatase I regulates nuclear gene expression in plants. Current Biology, 21(10), 897-903. doi:10.1016/j.cub.2011.04.004
- Wu, C., Suzuki, A., Washida, H., & Takaiwa, F. (1998). The GCN4 motif in a rice glutelin gene is essential for endosperm-specific gene expression and is activated by Opaque-2 in transgenic rice plants. The Plant Journal,14(6), 673-683. doi:10.1046/j.1365-313x.1998.00167.x
- Xu, R., Li, D., Li, H., Li, J., Yang, Y., Qin, R., … Yang, J. (2016). Isolation of four rice seed-specific promoters and evaluation of endosperm activity. Plant Cell, Tissue and Organ Culture (PCTOC), 128(1), 125-132. doi:10.1007/s11240-016-1091-5
- Ye, J., Yang, Y., Wei, X., Niu, X., Wang, S., Xu, Q., … Wang, S. (2018, April 19). PGL3 is required for chlorophyll synthesis and impacts leaf senescence in rice. Journal of Zhejiang University-SCIENCE B, 19(4), 263-273. doi:10.1631/jzus.b1700337
- Ye, X., Al-Babili, S., Kl?ti, A., Zhang, J., Lucca, P., Beyer, P., & Potrykus, I. (2000). Engineering the Provitamin A (?-Carotene) biosynthetic pathway into (carotenoid-free) rice endosperm. Science, 287(5451), 303-305. doi:10.1126/science.287.5451.303


