
Daleep Grewal
Roslyn High School
Round Hill Road
Roslyn Heights, NY 11577
Peer Reviewer: Himanshi Verma
Pro Reviewer: Dr. Madhav C Menon
Abstract
Chronic kidney disease prevalence in the United States is expected to rise substantially, indicating that kidney transplantation is sub-par and that it is necessary to find biomarkers of the disease. Shroom3 was identified as a gene whose expression in renal allografts correlates with renal function decline; this is significant as 40% of transplant patients were found to carry a Shroom3 mutation (Chuang & Menon, 2012). To study the role of the Shroom3 gene, Menon et al have used animal models to modulate Shroom3 levels; in podocytes, or specialized kidney cells, a protein-protein interaction was identified between Shroom3 and Fyn, another Src protein, which impacts podocyte actin cytoskeleton and cell size (Menon, 2018). It is imperative to elucidate whether Fyn has independent effects on podocyte morphology to delineate the mechanism of Shroom3, such that future therapeutics could target the protein. The present study manipulated Fyn in podocytes such that the protein was either inhibited, using an inhibitor, or knocked down via shRNA. Overall, podocyte actin organization was not affected upon FYN inhibition or Fyn knockdown. While there were increases in non-phosphorylated Src kinase fraction, no differences in podocyte cell size or phosphorylated Src were seen with inhibition. Upon Fyn knockdown, while no cytoskeletal changes were seen, podocyte cell size was reduced. These findings could suggest the importance of other Src family proteins in the function of FYN in its absence or after inhibition of activation. Future studies are required to modify the structure of the FYN inhibitor to confirm the effect of Fyn knockdown on podocytes. This study provides insight on cell signaling, regulation, and morphology, which can hopefully contribute to the discovery of new biomarkers that play a role in chronic kidney disease.
Introduction
Chronic kidney disease currently affects 30 million adults in the United States and is expected to rise substantially in the next 25 years (Chuang & Menon, 2012). The treatment of choice for chronic kidney disease is a kidney transplant; however, long-term kidney transplant outcomes currently remain sub-par, delineating an unmet medical need to detect signs of chronic kidney disease before it develops into a serious condition (McLaren & Menon, 2000). This study sought to interrogate the gene Fyn, which is prevalent in podocytes, specialized kidney cells responsible for glomerular function, and has potential for being a biomarker for chronic kidney disease development. This information may be useful in developing new therapeutics.
Genomic analysis most commonly entails identifying biomarkers, which are defined as measurable substances in an organism whose presence is indicative of phenomena such as disease, infection, or environmental exposure – in the context of this study, kidney disease (Redhead & Menon, 2017). Ideally, biomarkers should have: narrow biological variability, no influence by age or nutrition, and high sensitivity. Biomarkers can allow for early identification of kidney disease for patients at high risk of progression, and thus are urgently needed for early and targeted treatment to improve patient care (Rysz, Glubba & Francyzk, 2017).
It is important to delineate that this paper will be referencing both the gene and protein form of certain substances of interest, namely FYN and SHROOM3. When capitalized, the substance is being referred to as a protein (e.g. FYN), and when italicized, the substance is being referred to as a gene (e.g. Fyn).
Developments in genomics have led to a greater understanding of what plays a role in the development of kidney disease. Menon et al (2015) identified Shroom3 as a novel candidate gene whose expression in the renal allograft correlates with the decline of renal function, where 40% of kidney patients had an intronic Shroom3 mutation. This intronic locus is a modifier of Shroom3 expression, in people carrying A/A or A/G variants of this single nucleotide polymorphism (SNP). The ASD1-domain (the sequence motif that all SHROOM3 proteins are dependent upon) of SHROOM3 directly binds F-actin (the form of a protein that forms microfilaments) and thus could have direct independent morphologic effects on cell size. It has been observed that SHROOM3 knockdown of podocytes in animal models have smaller cell size in vivo and in vitro, decreased adhesion and increased motility (Yeo et al., 2015).
FYN is a member of the Src family of kinases typically associated with T-cell and neuronal signaling in development and normal cell physiology. Disruptions in these signaling pathways often have implications in the formation of a variety of cancers. By definition as a proto-oncogene, Fyn codes for proteins that help regulate cell growth. Changes in its DNA sequence transform it into an oncogene that leads to the formation of a different protein with implications for normal cell regulation (Ju et al., 2012). Through mRNA analysis FYN has been found to play a crucial role in synaptic function and plasticity, and central nervous system (CNS) myelination (Nygaard & Strimatter 2016).
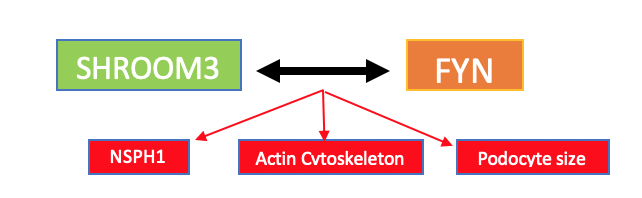
Menon and Murphy (2018) found a SHROOM3-FYN interaction via a novel SH3-binding domain that regulates FYN activation in podocytes, impacting the phosphorylation of NPHS1 (a gene that provides instructions for Nephrin, a protein necessary for the proper functioning of the renal filtration barrier), actin cytoskeleton and cell size. FYN-mediated phosphorylation of Nephrin, in tandem with Nck adapter proteins (signal-transducing), leads to actin filament assembly in podocytes (Verma et al., 2006); Figure 1 provides an illustrative representation this protein-protein interaction. FYN and other Src kinases are also known to be volume sensitive kinases, whose activation state is determined by cell volume alterations (Kapus et al., 2000). Therefore, the present study sought to determine whether these effects of Shroom3 on podocyte morphology and phenotype are Fyn-dependent. It was hypothesized that (1) there will be significant alteration in peripheral and perinuclear cytoskeletal integrity in Fyn knockdown cells, (2) inhibition of FYN will substantially damage podocyte cytoskeleton, and (3) the extent to which podocyte cytoskeleton will be altered will be greater in FYN inhibition cells compared to Fyn knockdown cells. To clarify, inhibition means that the level of FYN protein is completely wiped out in podocytes, while in knockdown, protein levels are only reduced.
Methods
Two groups were tested in this experiment: FYN inhibited and Fyn knockdown podocytes. An artificial inhibitor was designed by the Murphy Lab (Mount Sinai Hospital, Annenberg Building) to inhibit the function of FYN protein in selected podocytes. Using transfection techniques, plasmids containing Fyn gene knockdown material were injected into podocytes. Fyn knockdown podocytes were compared with Red Fluorescent Protein (RFP) Scramble podocytes that acted as a control; the control group was mutated at a random point in the cell’s DNA to ensure that the mutation itself was not causing changes to podocyte morphology. The following subsections explain the steps taken after the two groups of podocytes were created.
Western Blotting
Following a gel electrophoresis where a gel of different proteins was obtained, a transfer was conducted such that all the resolved proteins attached to a Polyvinylidene difluoride (PVDF) membrane. Following the transfer, the membrane was washed three times with Phosphate-buffered saline with Tween solution (PBST+). A primary antibody (isolated from rabbit) was applied overnight on a rocker in freezing temperature, followed by the secondary antibody (isolated by deer). The membrane was then washed three more times. Incubation with specific antibodies generated a chemiluminescence signal, indicating the amount of FYN protein within specific podocyte lines. The strength of the signal was compared with protein signals from RFP control podocytes in order to determine whether FYN manipulation was successful. See Figure 2 for an illustration of the western blotting process.
Cell Culture/ Sub-culturing
The cell lines for this experiment comprised of Fyn knockdown and FYN inhibition podocytes. The base medium for these cell lines was ATCC-formulated Dulbecco’s Modified Eagle’s Medium, Catalog No. 30-2002. To make the complete growth medium, fetal bovine serum was added to a final concentration of 10% to the base medium.
To continue the podocyte line, any culture media within the cell lines were removed and discarded. The cell layers were then briefly rinsed with 0.25% (w/v) Trypsin- 0.53 mM EDTA solution to remove all traces of serum and dead cells. Following that, 3.0 mL of Trypsin-EDTA solution was added to a 10 mL flask, kept under a lab hood. The cells were observed under an inverted microscope until cell layer dispersed (usually within 10 minutes). Afterwards, 8.0 mL of complete growth medium was added and the cells were aspirated by gentle pipetting. Finally, appropriate aliquots of the cell suspension were added to new culture vessels and were then incubated at 37°C.
Phalloidin (TRITC) Staining
Phalloidin is a bicyclic peptide that belongs to a family of toxins isolated from the Amanita phalloides mushroom. Phalloidin is commonly used in imaging applications to selectively label the F-actin of cell cytoskeletons in fixed cells; in the context of this experiment, it was used to examine the cytoskeleton of podocytes following FYN inhibition and Fyn knockdown.
Firstly, podocytes were fixed in collagen gels with 3.7% (v/v) paraformaldehyde for 10 min at room temperature. Afterwards the cells were rinsed 3 times in PBS. The blocking buffer (used to block non-specific binding sites) was then added; its composition was 2% BSA and Bovine serum albumin. Following the rinsing the podocytes were permeabilized with 0.1 %(v/v) Triton X-100 for 10 min at room temperature, and rinsed 3 more times in PBS. The cells were then rinsed with 0.165 ?M of Acti-stain Phalloidin for 30 min in the dark at room temperature and washed the cells 3 times with PBS. Subsequently, the stained samples were placed on glass slides and gently pressed with a glass coverslip using a drop of anti-fade mounting medium to prevent photo-bleaching. Finally, side of the coverslip was pressed with nail polish and stored the slides in the dark at 4°C.
Microscopy and Data Analysis
A fluorescence microscope (Zeiss Axiolmager) was used to examine the Phalloidin-stained slides. One hundred pictures were captured under the microscope of both Fyn knockdown and FYN inhibited cells. Fyn knockdown and/or inhibition was said to have an effect on actin cytoskeleton if at least 90 of the 100 pictures illustrated an alteration in size or structure of the cytoskeleton; it is common practice for most in vitro experiments to follow this model. The images captured were analyzed using Zen Pro software.
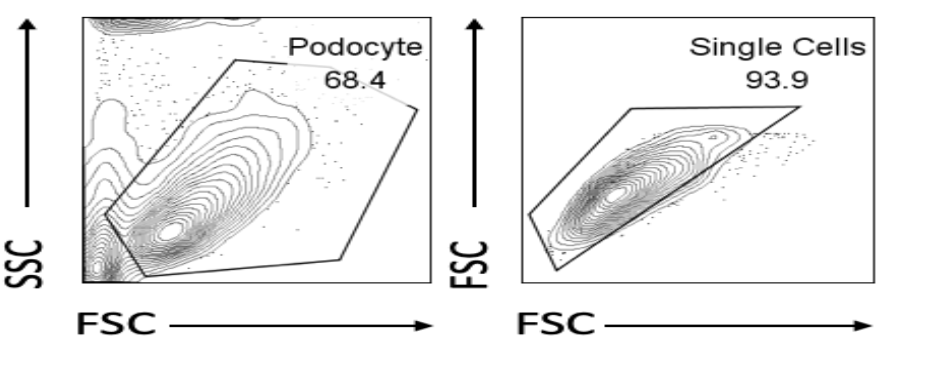
Flow Cytometry and Forward Scatter Estimation
Human podocytes (Fyn knockdown versus RFP) were incubated for 20 minutes at 4°C in either Annexin buffer (BD-556454) or PBS with 1:10 dilution of Annexin V Buffer (BD-550474) and 1:1000 viability dye (65–0863–14; eBioscience). As positive control, podocytes were pretreated with 25% w/v H2O2 for 20 minutes at 4°C before staining. Hydrogen peroxide is currently the most widely used apoptosis inducer; using this as a positive control is useful to ensure whether the cells being used in the experiment respond normally to substances known to induce specific cell responses.
Results
Western Blot
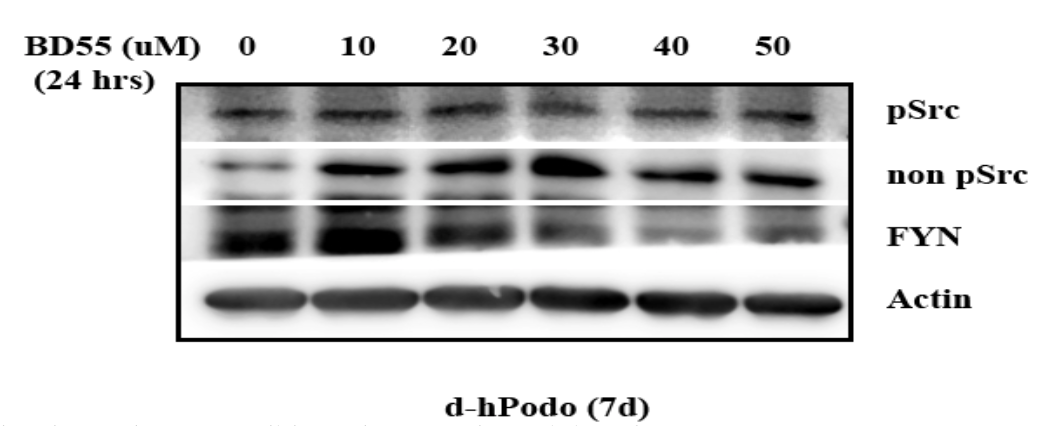
The first portion of this study sought to determine the effectiveness of the FYN inhibitor in order to test the second hypothesis that FYN inhibition will substantially damage podocyte cytoskeleton. Thus, a western blot, shown in Figure 3, was conducted to measure the amount of phosphorylated Src, non-phosphorylated Src, FYN, and Actin present in podocytes as the concentration of the inhibitor increased from 0µM to 50 µM. Figure 3 indicates that, although the amount of FYN decreased to a certain extent, the inhibitor did not decrease the activity of FYN as indicated by similar levels of phosphorylated levels of SRCs in untreated versus treated cell lysates.
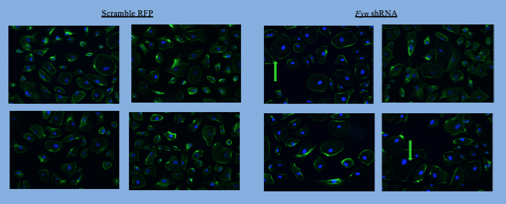
Phalloidin staining was used to examine the effect of FYN Inhibition and Fyn knockdown on podocyte cytoskeleton and size. Figure 4 shows that the actin organization of podocytes dramatically worsened as the concentration of the FYN inhibitor increased. Fyn knockdown podocytes appeared similar in size and overall shape to RFP control podocytes (Figure 5). Quantification of three experiments by different techniques showed a reduced size of Fyn knockdown podocytes versus control podocytes. Overall, the results support the third hypothesis in that FYN inhibition caused greater structural damage in podocytes than in Fyn knockdown podocytes.
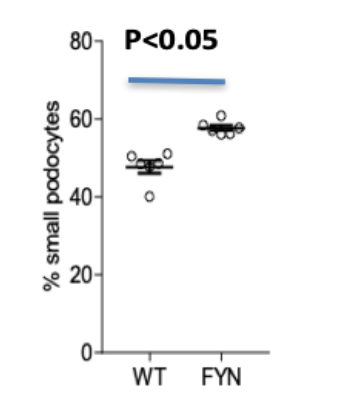
The flow cytometry experiment illustrates the overall shrinkage in podocyte size due to Fyn knockdown. Figure 6 and 7 illustrate the results of the flow cytometry experiment. Figure 6 identifies reduced forward-scatter mean fluorescence intensity, and Figure 7 indicates that Fyn knockdown podocytes comprised of a larger percentage of small podocytes compared to wild-type (RFP) podocytes.
Discussion
The present study sought to determine the role of Fyn in determining podocyte size and actin cytoskeleton by either inhibiting or knocking down the protein in selected podocytes. To examine this effect, Fyn was either knocked-down or inhibited and then examined using a Phalloidin-staining process.
As expected, the western blot showed the amount of FYN protein decreased as the concentration of the inhibitor increased. However, the overall pSrc amount remained relatively stable as the inhibitor increased in concentration (Figure 3). This indicates that the activity of FYN was overall unaltered, as the total phosphorylated (activated) amount of the Src-proteins remained the same.
To understand the physical effects of the inhibitor, I Phalloidin-stained the podocytes to examine their cytoskeletons. In accordance with the hypothesis, the podocytes lost cytoskeletal integrity as the concentration of the inhibitor increased; at 50 µM, cell death was observed as many podocytes had substantially shrunk in size (Figure 4). Thus, it can be asserted that FYN does have some independent effect on podocyte cytoskeleton and podocyte size. Upon further examination of the inhibitor, however, it was observed that the inhibitor was too toxic for podocytes, which serves as an additional explanation for why there was such a distortion in cytoskeletal integrity. Compounded with the results of the western blot, both pieces of the study’s results indicate that alterations to the inhibitor are needed such that the inhibitor drug can sufficiently wipe out the activity of FYN while still being compatible with podocytes such that it does not become too toxic for the cells.
The hypothesis that there will be a significant alteration in podocyte cytoskeleton as a result of Fyn knockdown was partially supported by the results. The images indicate that there was no overall difference, however some Fyn-knockdown podocytes were substantially smaller in overall cytoskeleton size as well as more circular compared to RFP Scramble podocytes, which were larger and more ovular in size (Figure 5). The flow-cytometry further illustrates this idea (Figures 6 & 7). Podocyte function depends on a highly ordered cellular arrangement of filtration compartments and correct signaling within their microenvironment. Thus, an increase in cytoskeletal size and alteration in overall shape can severely compromise their overall function, as they operate best when ovular and small such that they can collectively form a proper barrier within the kidney.
Various limitations were present in this study. For example, the inhibitor studies involved a series of dose dependent and time dependent course to conclude for drug efficiency. Additionally, the present study required terminally differentiated cells (7 days). Both of these burdens extended the time necessary to complete the experiments. Furthermore, the Phalloidin staining was occasionally unsuccessful, which led to multiple retries of experiment. As a result, the stain color was changed from red to green in order to obtain better images.
Conclusion
It is interesting to note that there were exceptions in both parts of the study showing a significant difference between the control and FYN/Fyn-manipulated podocytes, such that FYN-inhibition led to smaller cell size and Fyn-knockdown led to smaller cell size. Thus, further analysis is needed. The future extension for this study is to (a) alter the structure of the inhibitor and (b) increase the sample size of Fyn-knockdown podocytes in order to clarify this study’s results. This study demonstrated however that overall, the actin organization in podocytes is not affected upon inhibition treatment or FYN knockdown, which suggests the importance of other SRC family proteins to take over the function of FYN in its absence. This study brings forth new insight on podocyte function and will hopefully contribute to the discovery of new biomarkers that play a role in chronic kidney disease.
Acknowledgements:
My work was carried out during a summer internship under Dr. Madhav C Menon, MD, using research funding provided to him by the American Heart Association (grant 15SDG25870018) and by individual philanthropic support from Nina and Homer Eaton. All intellectual property associated with this work belongs to Dr Menon, who I deeply thank for providing me the opportunity to engage in such interesting research. Special thanks to Dr. Allyson Weseley (Roslyn High School) for reviewing and commenting on the contents of this study.
References
- Ju, W., Smith, S., Menon, M. (2012). Genomic biomarkers for chronic kidney disease. National Institutes of Health, 159(4),290-302, doi: 10.1016/j.trsl.2012.01.020
- Kapus, A., Ciano, C., Sun, J., Zhan, X., Kim, L., Wong, T., Rotstein, O. (2000). Cell volume-dependent phosphorylation of proteins of the cortical cytoskeleton and cell- cell contact sites. The role of Fyn and FER kinases. The Journal of biological chemistry 2(275), 32289- 32298. doi:10.1074/jbc.M003172200 ?
- McLaren, A., Menon, M. (2000). Chronic allograft failure in human renal transplantation. Journal of Kidney Research, 3(2), 98-103. doi:10.7228/JCI71380.
- Menon, M., Chuang, P., He, J. (2012). The glomerular filtration barrier: components and crosstalk. International Journal of Nephrology. 2012. 749010. 10.1155/2012/749010.
- Murphy, B., Menon, M. (2017). Novel therapeutics identification for fibrosis in renal allograft using integrative informatics approach. Science Reports, 7(3), 291-301. doi: 10.1038/srep39487
- Nygaard, H., Dyck, C., Strittmatter, S. (2014). Fyn kinas as a novel therapy for Alzheimer’s disease. Alzheimers Research Therapy, 6(1), 145-159, doi:10.1186/alzrt238.
- Readhead, B., Menon, M. (2017). The use of genomics and pathway analysis in our understanding and prediction of clinical renal transplant injury. Science Reports, 7(3), 291-301. doi: 10.1038/srep39487
- Rysz, J., Glubba, A., Francyzk, B. (2017). Novel biomarkers in the diagnosis of chronic kidney disease and the prediction of its outcome. International Journal of Molecular Sciences, 2(18), 1-17. doi: 10.3390/ijms18081702
- Verma, R., Kovari, I., Soofi, A., Nihalani, D., Patrie, K., Holtzman, L. (2006). Nephrin ectodomain engagement results in Src kinase activation, nephrin phosphorylation, Nck recruitment, and actin polymerization. The Journal of Clinical Investigation, 3(116), 1346-1359. doi:10.1172/JCI27414 ?
- Wei, C., Banu, K., Garzon, F., Basgen, J., Philippe, N., Yi, Z., Liu, R., Choudhuri, J., Fribourg, M., Liu T., Cumpelik, A., Wong, J., Khan, M., Das, B., Keung, K., Salem, F., Campbell, K., Kaufman, L., Cravedi, P., Zhang, W., O’Connell, P., He, J., Murphy, B., Menon, M. (2018) Shroom3-Fyn interaction regulates nephrin phosphorylation and affects albuminuria in allografts. Journal of the American Society of Nephrology, 29(11), 2641-2657, doi: 10.1681/ASN.2018060573 Yeo, C., O’Meara, C., Bonomo, J., Veth, K., Tomar, R., Flister, M., Drummond, I., Bowden D., Freedman, B., Lazar, J., Link, B., Jacob, H. (2015). Shroom3 contributes to the maintenance of the glomerular filtration barrier integrity. Genome Research, 2(25), 57-65, doi:10.1101/gr.182881.114.


