
NHSJS Reports
Thomas Barbera
Peer Reviewer: Sanya Jain
Professional Reviewer: Dr. Misty D. Smith
Abstract
Sudden Unexplained Death in Epileptic Patients (SUDEP) is one of the leading causes of epilepsy related deaths.The risk of SUDEP in children with Dravet syndrome is estimated to be 15-fold greater than other childhood-onset epilepsies (Skluzacek JV, Watts KP, Parsy O, Wical B, Camfield P. Dravet syndrome and parent associations: The IDEA League experience with comorbid conditions, mortality, management, adaptation, and grief. Epilepsia. 2011;52(suppl 2):95–101.) Research in this field is focused on discovering a biomarker, or way to predict, SUDEP. Research is focused on three possible biomarkers: respiratory rates, serum lactate, and neuroinflammation. The following experiments explore each possible biomarker in separate experiments. The first experiment explores respiratory rates following seizures, and the hypothesis is that declining respiratory rates cause respiratory failure during sleep. This experiment used a non-invasive respiratory collar to measure baseline respiratory rates and respiratory rates following hyperthermia-induced seizures. The second experiment studies serum lactate, and the hypothesis is that the increase in serum lactate following a seizure causes cardiac or respiratory failure during sleep, thus causing SUDEP to occur. This experiment used a lactate kit to analyze the concentration of serum lactate in a sample of blood collected following euthanization of the mice used in the respiratory rates experiment. The third experiment studies neuroinflammation of astrocytes, a specific neurotransmitter that controls the Nav1.1 Sodium Channel in the hindbrain, and the hypothesis is that the inflammation of the astrocytes following a seizure causes brain dysfunction, eventually resulting in SUDEP. This experiment analyzed brain slices using color-based imaging following the staining of the brain slices for specific proteins. For all three of the experiments, only the serum lactate study yielded significant results. The p-value obtained for the difference in lactate following a seizure was 0.04978, which indicates a significant difference as the value is >0.05. However, all other p-values obtained were at least 0.3168, which indicates none of the measured respiratory rates yielded a significant increase following seizures. Antiepileptic Drug (AED) production should be focused on controlling increase in serum lactate, and if the rate of SUDEP decreases for patients on this AED, a correlation between SUDEP and serum lactate increase can be established.
Introduction
Sudden Unexplained Death in Epileptic Patients (SUDEP) is the cause of death for over 20,000 epilepsy patients each year in the United States, and accounts for over 15% of all epilepsy-related deaths1. All epilepsy patients are at risk of SUDEP, although it mainly occurs in patients between 20 to 45 years old2. SUDEP most commonly occurs in Dravet Syndrome epilepsy, a genetic strain of epilepsy diagnosed in infancy3. SUDEP is nearly impossible to predict, as there have been no biomarkers validated as accurate ways to predict SUDEP. However, SUDEP is always preceded by seizures, so researchers agree that there must be validatable biomarkers to prevent SUDEP4. Mice bred for Dravet Syndrome exhibit similar characteristics to those of human patients, making mice viable options for testing for biomarkers to validate their development5. This leads researchers to question the pre-ictal (before a seizure) and post-ictal (after a seizure) rates in Dravet Syndrome mice, and whether they are significant enough to be classified as possible biomarkers for Sudden Unexplained Death in Epileptic Patients.
Literature Review:
Sudden Unexplained Death in Epileptic Patients (SUDEP)
Dravet Syndrome leads to an increased chance of SUDEP 6. SUDEP is classified as the death of an epilepsy patient that is both sudden and unexpected from an unknown cause. There does not have to be evidence of a seizure, and all examinations after the death of the patient must reveal no direct instigator of death7. Despite extensive studies on the causes of SUDEP and its frequency in Dravet Syndrome patients, there have been no completely validated biomarkers of SUDEP, making the deaths unpreventable and unpredictable8.
Some hypotheses as to what causes SUDEP are hindered respiratory, cardiac, and sensory functions following seizures; however, these causes are highly unpredictable, and there are no validated biomarkers for them9. Researchers discovered that generalized tonic-clonic seizures (GTCS), which involve the patient losing consciousness and experiencing severe muscle contractions, are a major risk factor for SUDEP10. One study found that SUDEP is 15 times more likely to occur in patients that have had more than two GTCS per month11.
Many experiments, both those aforementioned and several others, are investigating biomarkers for the prediction and prevention of SUDEP12. Along with conducting the pre-mortem (prior to euthanization) studies to discover biomarkers, researchers are attempting to expand their knowledge of SUDEP by performing post-mortem (following euthanization) analyses and autopsies with SUDEP patients. These studies are then compared to post-mortem studies of animal models and the differences are analyzed to obtain more information about the effects of SUDEP, and their correlations with animal models13.
Dravet Syndrome
Dravet Syndrome causes cognitive impairments such as slowed brain development, and is commonly accompanied by psychological issues and physical defects14. These psychological issues include anxiety and depression; furthermore, the mental defects that accompany Dravet Syndrome affect patients physically, as patients are more likely to develop unhealthy coping mechanisms such as smoking or develop an unhealthy diet.15. There are also specific characteristics that differentiate Dravet Syndrome from other forms of epilepsy14. Dravet Syndrome is a pharmacoresistant form of epilepsy, meaning AEDs do not affect the patient’s seizure frequency or amplitude. The syndrome is unique, as it is diagnosed when the patients are infants16.
Dravet Syndrome patients’ genetic mutation is commonly referred to as the SCN1A mutation, which is a mutation on the GABAergic neurons in the brain. GABAergic neurons are neurons that produce GABAergic receptors, which are inhibitory neurotransmitters (Ribak, C., 1987)). GABAergic receptors constrain the functions of sodium channels, which usually allow for communication between neurons through the transfer of sodium ions17. When these neurons produce damaged receptors, they can suddenly inhibit specific brain functions that cause the patient to seize18. Dravet Syndrome impairs the Nav 1.1 sodium channel, which directly affects the efficiency and strength of certain features of the brain in both the pre-ictal and post-ictal phases19.
Pre-Mortem Studies
Pre-mortem studies help researchers further understand SUDEP for more accurate predictions. Studies addressing pre-mortem biomarkers for SUDEP usually include investigating respiratory rates. Seizures in Dravet Syndrome are mainly caused by hyperthermia, commonly known as a fever. Hyperthermia-induced seizure tests begin by increasing the internal body temperature of the mouse, and simulating the thermal effect of a hyperthermia seizure20. The tests were previously administered to several age groups, and their conclusion supported the hypothesis that adults are more prone to SUDEP than children20.
Other pre-mortem studies focus on measuring respiratory rates, along with other breathing-related measurements such as oxygen saturation through both mouse assays and human testing. Sleep apnea, a sudden cessation of breathing during sleep, appears to have a connection to SUDEP21. Furthermore, several researchers hypothesize that tachycardia, an abnormally fast heart rate, occurs due to a decline in respiratory rates following seizures, thus causing SUDEP22. By studying the difference in pre-ictal and post-ictal respiratory rates, researchers may discover that hindered respiratory rates could be a possible biomarker of SUDEP.
Post-Mortem Studies
Some post-mortem studies explore peripheral lactate, a type of lactate produced when a body must perform a physically demanding action23. One study which analyzed the increase of lactate supported the conclusion that the increase of the UCH-L1 branch of lactate and plasma both correlate with age; however, the sex of the patient did not affect the levels of UCH-L1 or plasma, which fits with existing literature24. In another study exploring connections between general tonic-clonic seizures and SUDEP, the researchers discovered that serum lactate significantly increased after a GTCS compared to other forms of seizures. This significant difference in lactate between GTCS and other types of seizures presents the possibility of lactate as a biomarker to predict SUDEP25.
Some post-mortem tests examine the brains of Dravet Syndrome mice following their seizures. Astrocytes, nervous system sensors primarily found in the medulla that control breathing and heart rate, may send abnormal signals following seizures26. Astrocytes can also sense movements within the brain, specifically the changes in glucose and lactate, which activate the astrocytes and begin neuroinflammation27. Measuring astrocyte inflammation involves staining predetermined sections of the brain and imaging them28. The pictures are then analyzed for differences with saline-controlled slices through the staining and imaging systems to quantify differences in the pictures28.
One study focused on the correlation between epileptic seizures and non-epileptic seizures by monitoring the NaV1.1 sodium channel following seizures29. Researchers discovered that the greatest levels of dysfunction occurred following hyperthermia seizures29. This further suggested neuroinflammation as a possible biomarker for SUDEP. Another study discovered that over 50% of human patients tested had miniscule brain damage, and that approximately 25% of brains showed signs of swelling following SUDEP30. This study also found that there are no significant differences in brains due to age or sex in SUDEP cases, which helps researchers by allowing them to test a wider variety of brains without having to meet specific parameters30.
Methods:
Justification for Experiments
The literature on SUDEP and its possible biomarkers presents substantial evidence that respiratory, lactate, and neuroinflammatory analyses may reveal possible biomarkers for SUDEP. Due to the decrease in respiration following seizures, respiratory rates following seizures may be a possible biomarker for SUDEP22. The correlation between seizures and a positive increase in serum lactate following the seizures warrants an analysis of the levels of serum lactate in the body directly following the seizure, which may indicate an increased risk for SUDEP25. Because lactate directly affects neuroinflammation, inflammation of astrocytes in the medulla of the brain may also be a viable biomarker for SUDEP30. All of these potential biomarkers may lead researchers to further understand SUDEP and prevent it. This evidence and examination of literature suggests the hypotheses that differences between pre-ictal and post-ictal measurements in respiratory rates, serum lactate, and in Dravet Syndrome mice are substantial enough to provide accurate biomarkers for SUDEP.
Risk Assessment and Funding
All of the research was conducted in a University of Utah College of Pharmacy research lab and was supervised by qualified scientists. Qualified scientists supervised the use of all hazardous chemicals and all animals both during their uses and during their storage. All procedures were approved by the University’s Institutional Animal Care and Use Committee (IACUC) prior to experimentation. All researchers also underwent animal handler training prior to all experimentation and access to the animals. This training was sanctioned by the IACUC, provided by the University, and allowed for safe use of chemicals and animals during experimentation. Funding for the experiments comes directly from the University of Utah College of Pharmacy.
Subjects
The following experiments use mice diagnosed with Dravet Syndrome, as experiments that use these mice provide data that correlates almost indistinguishably with human Dravet Syndrome patients31. This provides reasons for the use of mice in these experiments instead of other smaller organisms, as other animals would not provide as accurate data as mice diagnosed with Dravet Syndrome. The experiments used the same 12 mice, 6 with the SCN1A mutation (with Dravet Syndrome) and 6 from the same parents but without the mutation (Wild Type). These values were chosen to ensure that data would be conclusive without extending the research beyond the length of time available. The mice were between the ages of 2-4 weeks old and were obtained from a University breeding center for Dravet Syndrome mice. During experimentation, the mice were given arbitrary numbers by a third party to prevent bias throughout the study.
Respiratory Study:
Hyperthermia-Induced Seizure Induction
Hyperthermia seizures were induced before collecting post-ictal measurements of respiratory rates. A thermometer was inserted rectally into the mouse to allow for measurement of the internal body temperature of the mouse while it was heated. The seizure was then induced by placing each mouse into individual containers with a heat lamp placed above each container. Each mouse was monitored until it either seized or reached a temperature of 42.5?, as previous studies show that Dravet Syndrome mice must seize before reaching this temperature32. If the mouse seized, the temperature at which this happened was recorded and the mouse was immediately removed from the container and moved back to continue respiratory data collection.
Respiratory Testing

A respiratory collar was used to record all respiratory rates to analyze during data analysis. The mice had their necks shaved prior to the collar testing, and were then habituated to a new area for at least 30 minutes. The first mouse was placed into a testing container, and a practice collar (figure 1) was placed on the mouse for five minutes to acclimate the mouse to the collar before beginning the test.

The test collar (figure 2) was then placed around the neck of the mouse. Data was then recorded using the program MouseOX, which is a non-invasive monitoring system designed specifically to record respiratory functions of mice using a collar placed around the neck of the mouse33. After 5 minutes of collecting data, the collar was then taken off of the mouse, and the mouse was moved to another section of the lab to have a hyperthermia seizure induced. After the hyperthermia seizure was induced, the mouse was immediately returned to the test collar to have its post-ictal respiratory rates measured. After pre-ictal and post-ictal measurements were taken, the data was saved onto the computer and exported.
Serum Lactate Study:
Serum lactate was collected and run through a lactate kit to determine if there was an increase of lactate following a seizure. Each mouse was perfused and had their trunk blood, cerebral blood, and brain collected to obtain the materials for the lactate and neuroinflammation studies. Each of these blood samples were taken and mixed with a lactate kit to allow for spectrophotometric analysis: analysis done by shining a light through a solution to determine concentration (Lundgaard, I et. al, 2017). To allow for a standard curve during data analysis, 10 µL of Lactate Standard (LS) was mixed into 990 µL of Lactate Assay Buffer (LAB) and stirred together. Each well was then filled with 10 µL of the solution, and more LAB was added to obtain molarities ranging from 2-10 µM of the Lactate Standard. Samples of lactate taken from both Dravet Syndrome and wild type mice were then placed into each of the wells containing the solution.
Test samples were created to convert from absorption to lactate using the standard curve to analyze the difference in lactate following a seizure. In a separate well container, each lactate test sample was placed in a well with 50 microliters (50 µL) of a solution containing 46 µL of LAB, 2 µL of Lactate Substrate Mix (LSM), and 2 µL of Lactate Enzyme Mix (LEM). After the serum was diluted in the solution for two hours, samples from the wells were then moved to different wells containing the original 50 µL solution to obtain dilution values closer to the standard curve. The solutions were then incubated in the new dilution for 30 minutes before spectrophotometric analysis began.

Solutions then had to be run through a spectrophotometer to collect data on the amount of lactate in each sample. Both the standard curve solutions and the test solutions were then placed into an ELISA spectrophotometer (figure 3). This specific spectrophotometer is designed to measure levels of lactate in blood samples34. The values collected from the spectrophotometer were then exported from the computer and later used for data analysis.
Neuroinflammation Study:
Brain Slicing

A brain sample was taken from each mouse during their perfusion, and the hind portions were sliced to allow for staining of astrocytes. The slicing process began by placing an Optimal Cutting Temperature (O.C.T.) compound on a microtome (figure 4) at -20.0 °C. Immediately after the compound was applied to the surface, the brain was placed into the compound. A 40 micrometer (40 µm) slice was then taken from the brain. The slice was then taken off of the blade and was placed into a well filled with 1X PBS (phosphate-buffered saline) (see figure 5) if it contained the medulla of the brain. The well was then marked with the designated section of the brain that the slice was from. This process was continued for each brain, and then the slices were mounted.

Brain Mounting
Brain slices were then moved to image slides to allow for staining, photography, and quantitative analysis. After all of the brain slices were put into wells, the slices were mounted onto Superfrost Plus slides (figure 6). Each slice was mounted using 1X PBS and a paintbrush, and each slide contained 3-4 slices. Gel from an Immuno Edge pen was then applied around each slice to prevent the solutions from sliding off of the slides. After the gel was applied, the gel was allowed to dry until it was a translucent ring. The slides were then washed for five minutes three times and then moved into a humidified black slide box in preparation for their staining.

A solution composed of 1% Sodium Deoxycholate, 0.2% Triton X-100 in PBS was placed on each of the slices, and the slices were kept in the humidified box at room temperature while the solution permeated the slices35. The solution was then suctioned off of each slice. After the solution was suctioned off of the slices, the glial fibrillary acidic protein (GFAP) antibody, which was conjugated to Cy3 (Sigma) and diluted to 1:1000 in Cyto Q and Sodium Deoxycholate, was placed onto each of the slices and they were incubated in the antibody for two hours. The GFAP antibody stains for GFAP, which appears on astrocytes. The antibody was then removed via the suction method, and the slices were washed35. A 4′,6-diamidino-2-phenylindole (DAPI) solution diluted in 1X PBS was then placed onto each of the slices and they were incubated for 10 minutes in the solution. The sections were washed again before coverslips were applied using a Prolong Gold (Invitrogen) solution and placed into non-humidified slide boxes stored at 4? until they were viewed and photographed using the imaging microscope.
Photography

Each of the slides were taken and placed individually under the microscope (figure 7). GFAP staining appears under red light and DAPI staining appears under blue light, so both a red and a blue light were shined onto each of the slides separately to scan for levels of GFAP and DAPI respectively. Photographs were taken of several sections on each slide under each color28. Each photograph was also marked with the arbitrary number given by the third party that correlated with the mouse whose brain slices were on each slide.
Data Analysis/Results:
Respiratory Study
A significant difference between post-ictal and pre-ictal respiratory rates may further validate the use of respiratory rates as a biomarker for SUDEP, as respiratory complications may cause SUDEP to occur. A p-value obtained from a t-test quantifies a difference between two data sets as either significant or insignificant based on the p-value obtained from the test. The p-value obtained must be below or equal to 0.05 to be classified as a significant difference between the data sets36. The standard deviation of the data sets shows whether the range of the data set varies widely from the mean of the data. This helps classify the data as accurate or inaccurate, as a low standard deviation compared to the mean value means the data points are all similar in value.
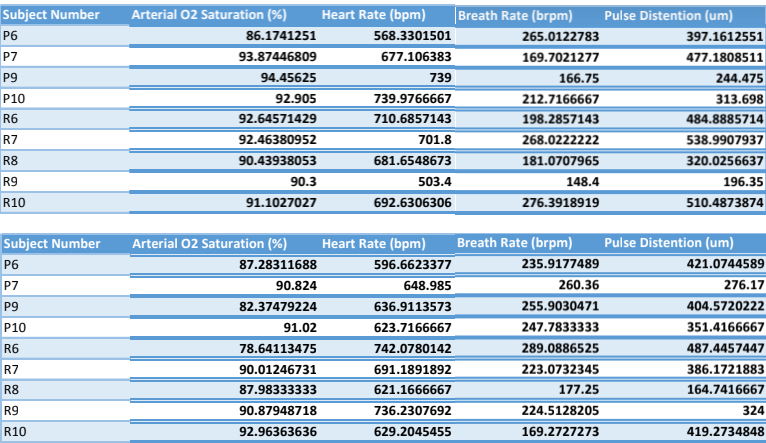
Figure 8a: Tables of pre-ictal and post-ictal measurements of oxygen saturation, heart rate, breath rate, and pulse distension of the Dravet Syndrome (P 1-4) and Wild Type (R 1-5) mice
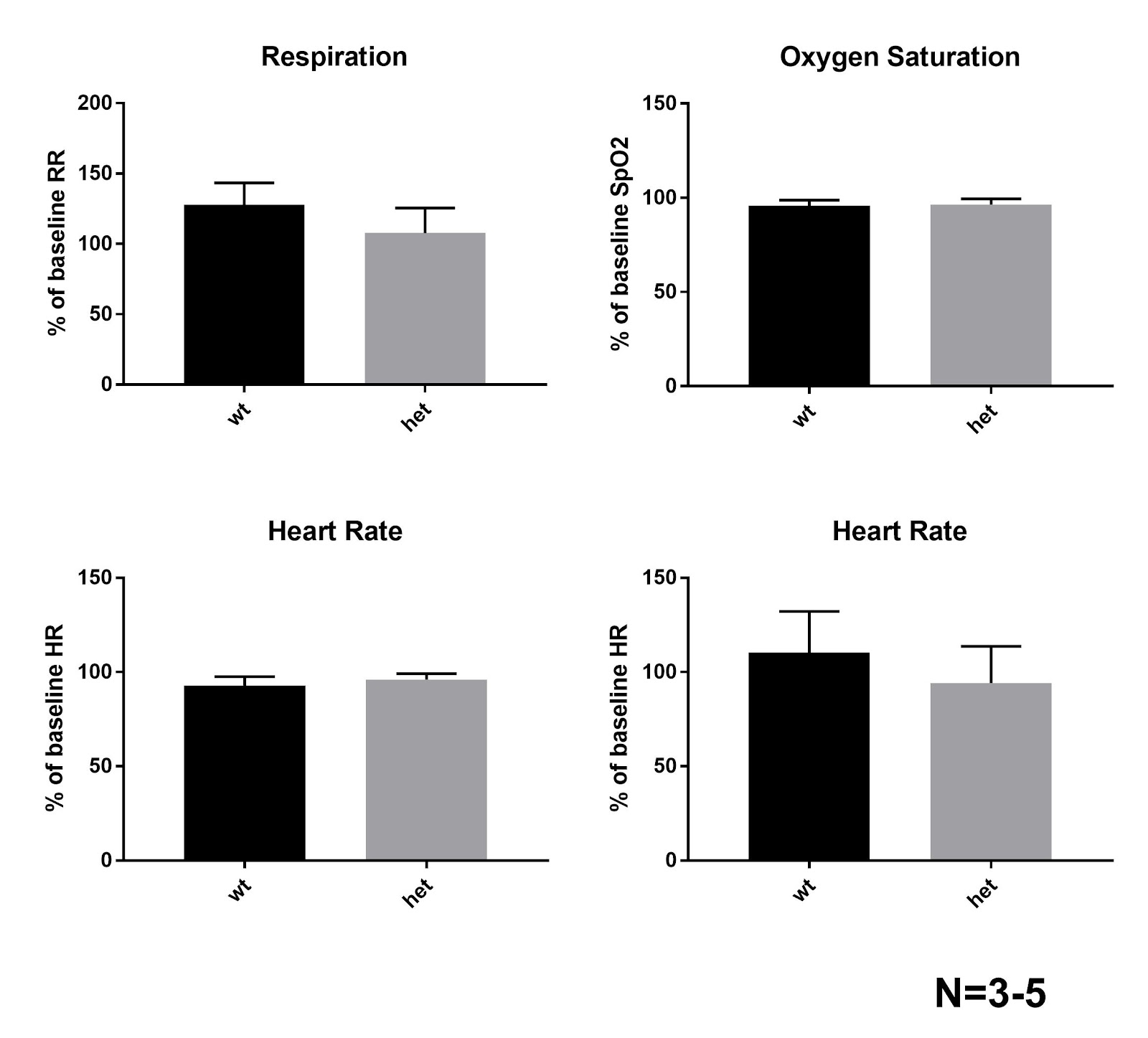
Figure 8b: (Beginning in upper left corner) Graphs of respiration, oxygen saturation, (SpO2) heart rate, and pulse distension of wild type (wt) and heterozygous (het) mice. The standard deviation of each mean is shown with a line above the mean value (apex of the grey region). All t-tests show no significance in the difference between respiratory rates following seizures compared to pre-seizure rates (p> 0.05)
The average of the differences for each of the four measurements of the five Dravet Syndrome and wild type mice were obtained from the MouseOX program (figures 8a & 8b). The standard deviation of the data from each of the measurements is shown using a line beginning at the mean value to the positive standard deviation (figure 8b). The averages for oxygen saturation of the wild type and Dravet Syndrome mice are 95.80±6.09% and 96.44±6.79% of the baseline oxygen saturation respectively with a p-value of 0.8878. The averages for the heart rate of the wild type and Dravet Syndrome mice are 92.82±9.56% and 106.2±23.1% of the baseline respectively with a p-value of 0.3168. The averages for respiration (breath rate) of the wild type and Dravet Syndrome mice are 128.1±31.3% and 107.9±39.3% of the baseline respectively with a p-value of 0.4324. The averages for pulse distension of the wild type and Dravet Syndrome mice are 100.8±21.2% and 86.14±27.74% respectively with a p-value of 0.5482. None of these p-values proved the difference in ranges significant, as none of the values are below 0.05. Furthermore, none of the standard deviations vary widely from the mean, so the data is statistically precise.
Serum Lactate Study
Lactate is created following a seizure due to muscle contractions in the heart, and it is hypothesized that a significant increase in lactate following a seizure could be a possible biomarker for SUDEP. The data obtained from this experiment was used to create a standard curve of lactate with respect to the optical density (O.D.) to convert the absorptions of the test samples to micromolarity (figures 11 a&b).
| Sample | Concentration | Found Conc. | Wells | Values | MeanValue | Std.Dev. | CV% |
| Sta01 | 2 | 2.691 | C1 | 1.1 | 1.1 | 0 | 0 |
| Sta02 | 2 | 2.691 | D1 | 1.1 | 1.1 | 0 | 0 |
| Sta03 | 4 | 4.651 | E1 | 1.508 | 1.508 | 0 | 0 |
| Sta04 | 4 | 4.055 | F1 | 1.384 | 1.384 | 0 | 0 |
| Sta05 | 0 | -0.552 | G1 | 0.425 | 0.425 | 0 | 0 |
| Sta06 | 0 | -0.605 | H1 | 0.414 | 0.414 | 0 | 0 |
| Sta07 | 6 | 4.92 | A2 | 1.564 | 1.564 | 0 | 0 |
| Sta08 | 6 | 6.149 | B2 | 1.82 | 1.82 | 0 | 0 |
| Sta09 | 8 | Masked | C2 | Masked | Masked | Masked | Masked |
| Sta10 | 8 | Masked | D2 | Masked | Masked | Masked | Masked |
Figure 9: Concentration of each well obtained by the spectrophotometer with their respective means, standard deviations, and coefficient of variation (%CV)
| hets | nmol | dilution factor | calc (uM) | |
| 1R | 0.04635522997 | 100 | 4.635522997 | 0.4635522997 |
| 2R | 1.007085385 | 100 | 100.7085385 | 10.07085385 |
| 4R | 0.3201633241 | 100 | 32.01633241 | 3.201633241 |
| 5R | 0.6444097514 | 100 | 64.44097514 | 6.444097514 |
| wt | ||||
| 1P | 0.03914975381 | 100 | 3.914975381 | 0.3914975381 |
| 2P | 0.3585925303 | 100 | 35.85925303 | 3.585925303 |
| 3P | -0.220247388 | 100 | 0 | 0 |
| 4P | -0.2538729434 | 100 | 0 | 0 |
| 5P | -0.1722108803 | 100 | 0 | 0 |
Figure 10: The nanomoles, dilution factor, and molarity for the Heterozygous (1R-5R) and Wild Type (1P-5P) mice
The absorptions obtained from the ELISA spectrophotometer for the test samples were then converted to nanomolarity using the line of best fit on the standard curve. The r2 value on the line of best fit is 0.95, indicating a linear relationship between molarity and wavelength. This value statistically indicates the line of best fit is an accurate way to convert wavelength to molarity, the amount of lactate per liter of blood. All of the wavelengths obtained for the test solutions were then converted to molarity by using the equation for the line of best fit (figure 11a). The mean micromolarity (µM) of the lactate for the wild type mice was 0.1500±0.2057 µM. The mean micromolar lactate for the mice with Dravet Syndrome was 5.045±4.147 µM. A t-test analysis on the difference between the micromolarity of the samples from the Dravet Syndrome and wild type mice yields a p-value of 0.04978, indicating a significant difference between the lactate created by the Dravet Syndrome mice and the lactate created by the wild type mice.
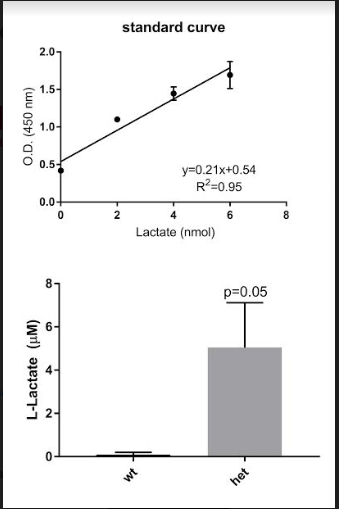
Figure 11a (Top): The standard curve created from the base solutions of LS and LAB. The optical density (O.D.) of 450 nm was specifically selected, as it allows for minimum absorption of chemicals from the lactate kit. Equation in upper right-hand corner was used to convert test samples from absorption to molarity.
Figure 11b (Bottom): Graphs of micromolar lactate of the wild type (wt) and heterozygous (het) mice following the conversion from O.D. to micromolarity. P-value showed significance (p?0.05) between post- and pre-ictal rates.
Neuroinflammation Study
Astrocytes inflame following a seizure, and it is hypothesized that this inflammation inhibits their ability to produce proteins. If this production is hindered, this will cause complications in the Nav1.1 Sodium Channel, which relies on these proteins, making neuroinflammation a possible biomarker for SUDEP warrant SUDEP.
The quantitative differences between the photographs taken of the DAPI solutions were negligible. 3-Dimensional quantitative analysis was unable to determine any difference between the Dravet Syndrome and wild type slices. Because DAPI production is not hindered following inflammation, the protein must be contained rather than produced by astrocytes.
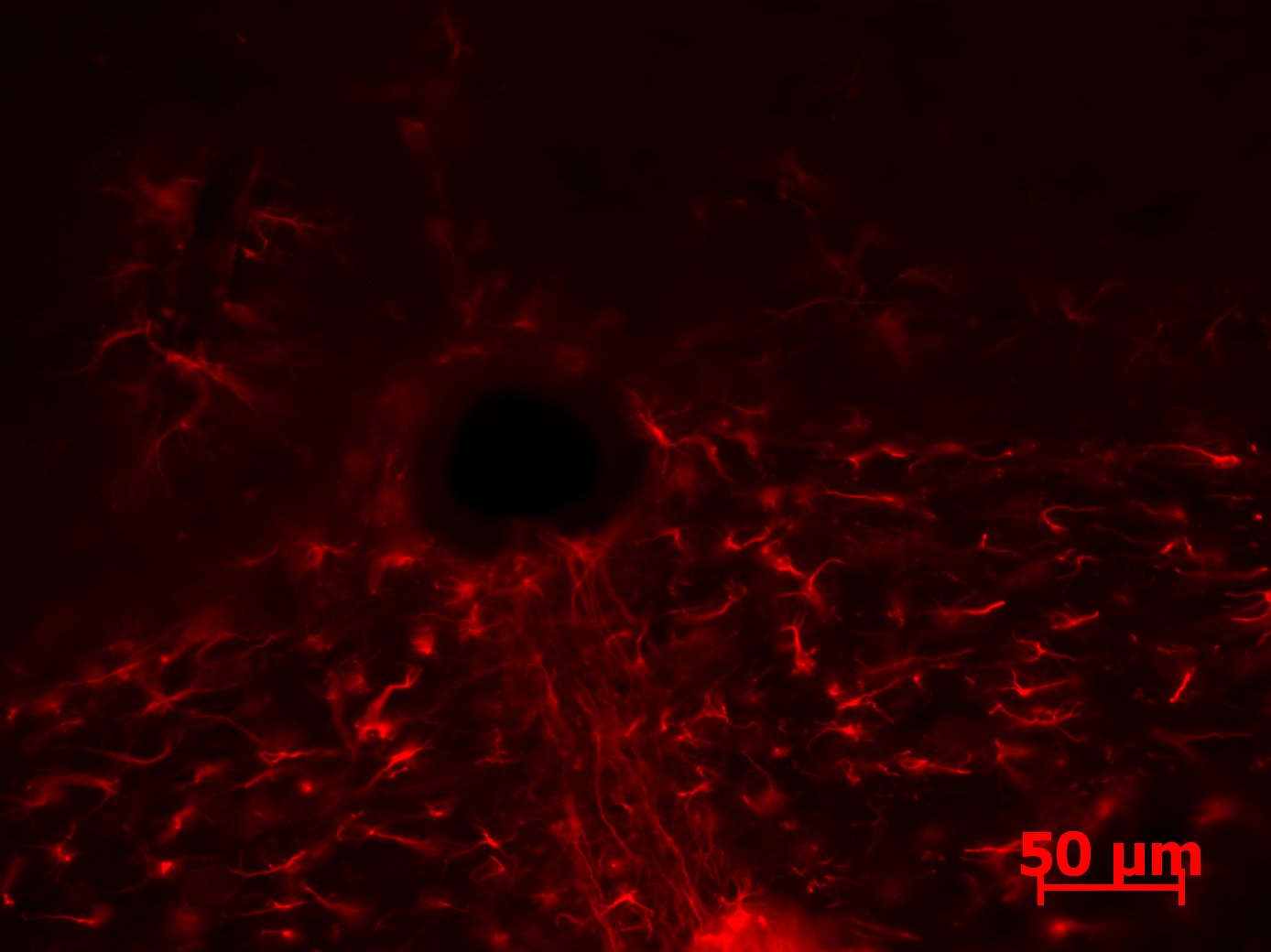
Figure 12: Photograph of the GFAP staining on the hind brain of a Dravet Syndrome mouse. Astrocytes are outlined in red, with the inflamed astrocytes appearing much larger with enlarged shapes. Brain shows much more inflammation than the Wild Type brain.
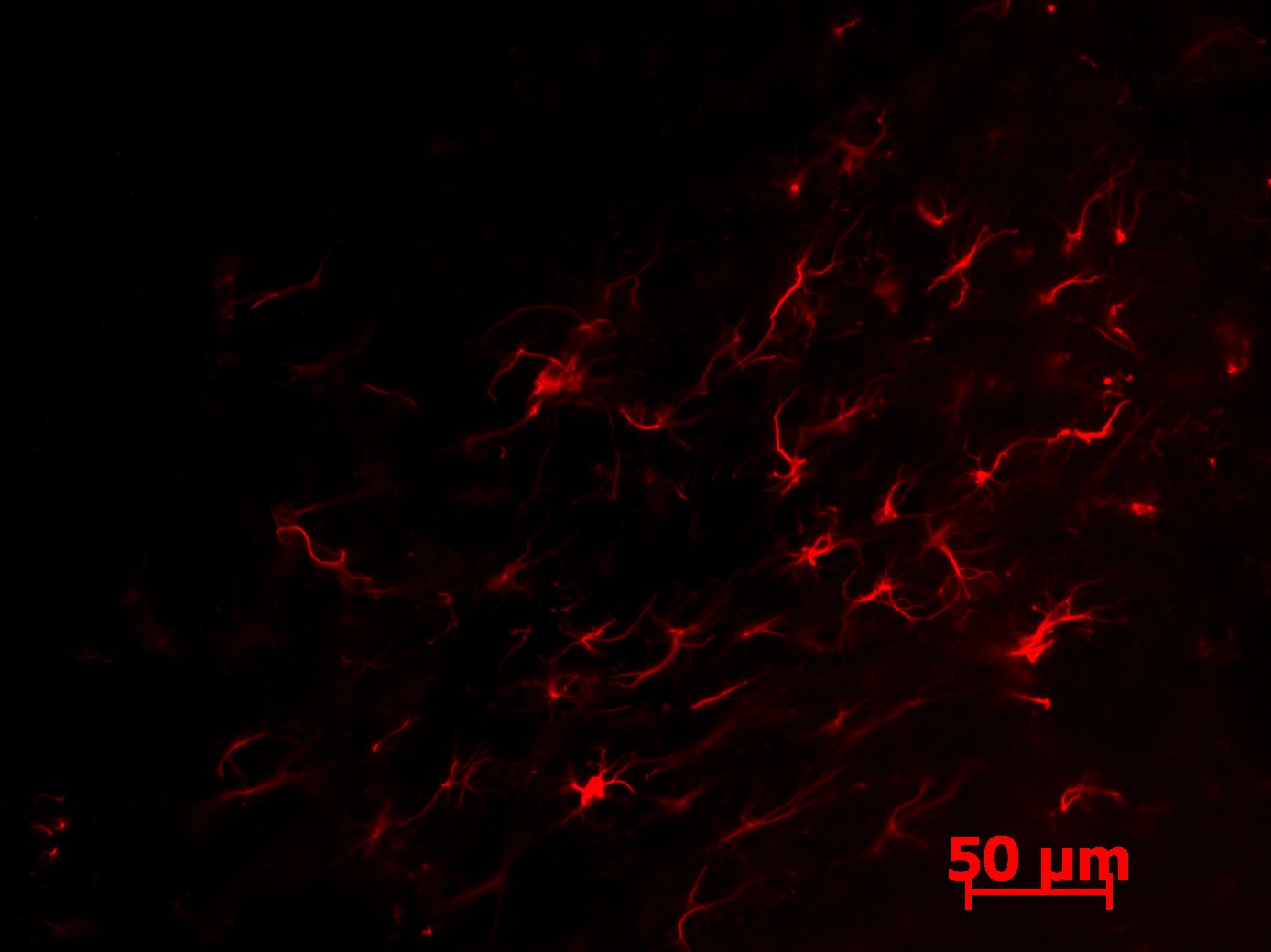
Figure 13: Photograph of the GFAP staining on the hind brain of a Wild Type mouse. Astrocytes are outlined in red, with the inflamed astrocytes appearing much larger with enlarged shapes. Brain shows much less inflammation than the Dravet Syndrome brain.
The images for the GFAP-stained slices showed more promising results. Inflamed astrocytes appear in jagged lines or are more enlarged shapes. The Dravet Syndrome image (figure 12) shows much more inflammation than the wild type image (figure 13). These images are potential indicators of the difference in inflamed astrocytes following a seizure; however, these images provide no quantitative data. 3-Dimensional quantification must be done to allow for analysis of the number of inflamed astrocytes, as astrocytes may appear on other layers of the slices or may be vertical in the slice and therefore are not captured by the image.
Conclusions and Discussion
Respiratory Rate Study:
The hypothesis for this study was not supported by the data. All data from this experiment yields no significant difference between any of the respiratory rates when comparing the post-ictal and pre-ictal measurements. The data was highly varied, as both respiration and pulse distension were lowered following seizures, whereas oxygen saturation and heart rate were increased following seizures, further supporting the conclusion that respiration is not an accurate biomarker for SUDEP. When compared, mean values for wild type mice versus Dravet Syndrome mice showed no significant difference, as p-values were at least 0.3168, indicating all differences were insignificant. Furthermore, standard deviations do not widely vary from the mean values, as the values were on average ±20.635%, which further supports the insignificant results obtained from the t-tests. This data effectively closes the avenue of research investigating respiratory rates as a biomarker for SUDEP.
Limitations for this portion of the experiment mainly appear during the hyperthermia-seizure induction. When mice were moved between the respiratory collar and the hyperthermia container, handling-induced seizures would sometimes occur. To minimize handling time, the container was moved directly next to the collar. Another limitation to this experiment was the respiratory collar data measurements. If the mouse moved during the data collection, the MouseOX program would fail to record specific data values. To minimize the effect on the data, the program gave error code. Every data point with an error code was omitted from the data analysis to prevent any inaccurate readings from the data analysis to provide the most accurate analysis from the data collected.
Serum Lactate Study:
This data from this experiment yields the most significant difference after analysis, and therefore correlates with the hypothesis that serum lactate significantly increases following a seizure. The molarities for the test samples are accurate, as the r2 value obtained from the line of best fit provides an accurate linear equation to convert the measurement obtained from the spectrophotometer to molarity. The p-value for the difference between the post-ictal rates for the wild type and Dravet Syndrome mice is 0.04978, proving the difference significant. This statistically significant difference offers substantial evidence to further this avenue of research. The hypothesis that lactate would significantly increase following seizures was supported by the data. Therefore, the large increase of lactate following seizures has the greatest potential of being a biomarker for SUDEP. As this experiment solely proves lactate increase following seizures, further experiments must determine whether there is a connection between this increase in lactate and SUDEP.
However, as the significance value is close to 0.05, limitations are greatly important, as they may cause the difference to become insignificant. Furthermore, the standard deviation of the experiments is at least ±82.20% of the mean value, indicating varied data, which further emphasizes the significance of the limitations. The main limitation to this experiment is the spectrophotometric analysis of other forms of lactate. Lactate is mainly produced when muscles contract, and during a seizure, specifically a tonic-clonic seizure, the patients must have extensive muscle contractions. Serum lactate is only produced by muscles in the heart during a muscle contraction. Other forms of lactate may be incorrectly analyzed as serum lactate during spectrophotometric analysis, and therefore obfuscate the final results. To minimize this error, each solution was diluted twice using the lactate kits. Because the kits are specifically designed to only allow pigments of serum lactate, this minimized the amount of other forms of lactate collected by the spectrophotometer.
Neuroinflammation Study:
The images produced from this experiment offer promising results for a 3-dimensional quantification of GFAP production following neuroinflammation. However, the lack of data to analyze from this experiment is due to the main limitation of the experiment itself: the 3-dimensional quantification of GFAP. Technology is available to quantify proteins such as the DAPI protein. However, there is still no technology available to quantify the amount of GFAP produced by inflamed astrocytes. This prevents a plausible correlation between the hypothesis for this experiment and the slices obtained from the experimental process. Technology, such as the
instrument used to analyze DAPI staining, is currently being refined to allow for GFAP quantification. Once this technology is refined, the quantification will be possible, and therefore the hypothesis can be tested against the data obtained from the quantification of the slices and their images.
Another limitation for this experiment is the limited data due to a limited sample size. Several hind brains were deformed or defiled during brain slicing, and therefore could not be used for staining. This was minimized by freezing the brains several ways before slicing began, and freezing them to both the O.C.T. compound and the Microtome prior to slicing. During photography, several slices appeared to have only regions stained rather than the entire slice, so these slices were omitted from data analysis. These limitations do not alter the data collected from the remaining slices; however, they further limit the amount of data that can be used to compare the inflammation in the Dravet Syndrome brains versus the wild type brains.
Further Directions
The initial research question focused on furthering potential biomarkers for predicting SUDEP to allow for the creation of an AED that could control the biomarker and therefore decrease the rate of SUDEP. Serum lactate had a significant difference following a seizure, thus answering the initial research question, as serum lactate increase proved to be the only potential biomarker according to currently analyzed data. Therefore, serum lactate is the only potential biomarker that warrants further directions. The first direction that must be pursued in this avenue of research is the expansion of the test group. An exact or extremely similar study should be performed with a larger test group, as the data from the original experiment provided a borderline significance. Further research must also substantiate that any seizure phenotype not occuring due to Dravet Syndrome would not cause similar changes in lactate. If the data maintains its significance following an increased number of subjects, and can determine the increase in lactate only increases following Dravet Syndrome seizures, the research can be used for other studies involving SUDEP.
This research presents data that narrows the creation of antiepileptic drugs (AEDs) to focus on preventing the increase in lactate following seizures. As the increase in lactate may cause cardiovascular or respiratory complications, maintaining a similar level of serum lactate post-seizure will allow for a larger understanding of SUDEP. If SUDEP rates decrease when patients are taking the AED, then a correlation can be established between an increase of serum lactate and the occurrence of SUDEP. If the rate of SUDEP does not decline after an AED has been created to decrease serum lactate following seizures, other hypotheses as to what may cause SUDEP to occur should be studied. However, as the other two avenues that were explored did not provide promising results, focusing AED production on controlling the increase in serum lactate following a seizure appears to be the most promising avenue of research to establish a biomarker of SUDEP.
Currently, further experiments are being conducted to attempt a correlation between the increase in lactate and SUDEP. As the study previously conducted solely discovered a significant increase in serum lactate, further experiments must prove the correlation between this increase in serum lactate and SUDEP. The current hypothesis for further research based off of the serum lactate data presented in this paper is that lactate following seizures may change more dramatically if the patient is susceptible to SUDEP. Researchers are currently monitoring lactate increase following a single seizure induced using hyperthermia-induction. The researchers are monitoring the occurrence of SUDEP in Dravet Syndrome mice while simultaneously having their serum lactate levels measured to determine whether there is a correlation between a lasting increase in serum lactate following seizures and SUDEP. If there is a significant correlation between a lasting increase in serum lactate and SUDEP, a correlation can be established between serum lactate levels following seizures and SUDEP, therefore making serum lactate a biomarker for Sudden Unexplained Death in Epileptic Patients.
Works Cited
- Gano & Grabenstatter, 2017 and Devinsky, Orrin, et al., 2016 [↩]
- United States Morbidity and Mortality Weekly Report, 2010, Devinsky, Orrin, et al., 2016, and Gano & Grabenstatter, 2017 [↩]
- Skluzacek JV, Watts KP, Parsy O, Wical B, Camfield P. Dravet syndrome and parent associations: The IDEA League experience with comorbid conditions, mortality, management, adaptation, and grief. Epilepsia. 2011;52(suppl 2):95–101. [↩]
- Lhatoo, Samden, et al. 2015, and Metcalf, Cameron et al. 2018 [↩]
- Metcalf, Cameron et al. 2018 [↩]
- Cheah, et al, 2012 [↩]
- Devinsky, Orrin, et al., 2016 [↩]
- Lhatoo, Samden, et al., 2015 [↩]
- Gano, L. B., & Grabenstatter, H. L, 2017 and Bozorgi, A., & Lhatoo, S. D. 2013 [↩]
- Harden, C., et al. 2017 [↩]
- Harden, C., et al. 2017 and Bozorgi, A., & Lhatoo, S. D. 2013 [↩]
- Bozorgi, A., & Lhatoo, S. D. 2013 [↩]
- Velagapudi, P. et al, 2012 [↩]
- Volkers, L. et al, 2011 [↩] [↩]
- Kobau, R. et al, 2008 [↩]
- Ceah, C. et al, 2012 [↩]
- Sugawara, T. et al, 2001 [↩]
- Ribak, C., 1987 and Oakley, J et al. 2009 [↩]
- Parihar, R. and Ganesh, S., 2013 [↩]
- Oakley, J. et al, 2009 [↩] [↩]
- Kim, Y., et al. 2018 [↩]
- Metcalf, Cameron et al. 2018 and Kim, Y., et al. 2018 [↩] [↩]
- Metcalf, C. S., et al. 2018 [↩]
- Mondello, S., et al. 2012, Oakley, J. et al, 2009 [↩]
- Matz, O., et al. 2016 [↩] [↩]
- Fuller S. et al, 2012, Metcalf, C. S., et al. 2018, and D., Accorsi, et al. 2013 [↩]
- MK, Jha, et al. 2016 [↩]
- Brackenbury, W., et al. 2013 [↩] [↩] [↩]
- Yamakawa, K., 2001 [↩] [↩]
- Thom, M., et al. 2016 [↩] [↩] [↩]
- Metcalf, Cameron et al. 2018 and Cheah, C. et al, 2012 [↩]
- Volkers, L., et al, 2011 [↩]
- Bittolo et al., 2016 [↩]
- Lundgaard, I et. al, 2017 [↩]
- Takahashi et al, 2016 [↩] [↩]
- Nurminen, M., 1997 [↩]


