
The Effect of Exercise on Regulatory T Cell Populations in Rat Mammary Adenocarcinoma Tumor and Spleen Samples using Foxp3 and CD25
Sharonya Battula
ABSTRACT
In healthy individuals, regulatory T cells (Tregs) suppress self-reactive lymphocytes to prevent autoimmune disease; however, in cancer patients, Tregs also serve a protumorigenic function, as their presence in the tumor microenvironment interferes with the ability of other lymphocytes to target and eliminate cancer cells. Because the progression of cancer is largely determined by the interactions between particular lymphocytes and cancer cells, higher concentrations of Tregs correspond with shorter lifespans among cancer patients after diagnosis.
Evidence shows that the removal of Tregs can enhance the anti-tumor immune response. Moreover, the depletion of Tregs is even considered an effective cancer immunotherapy treatment strategy for certain cancers. Other forms of treatment include exercise, which has been reported to reduce the risk of cancer recurrence and mortality, as well as regulate and deter the growth of tumors. Because of the effects reported from these treatment methods, the notion that exercise indirectly combats cancer by influencing specific lymphocytes became an idea of interest.
This research is concerned with the effect of exercise on regulatory T cell populations in the tumors and spleens of tumor-bearing rats. It was hypothesized that exercise decreases the proliferation of Treg cells in the tumor microenvironment and lymphoid tissue. For this study, tumor and spleen samples were collected from sedentary and exercised cohorts of rats with breast cancer. Subsequently, CD25 antibody, Foxp3 antibody, and Hoechst dye were applied to the tissues so that samples could be visualized using fluorescence microscopy to identify Tregs. Overall trends in the data suggest that exercise reduces Treg cell count in both the tumors and spleens of individuals, thereby limiting the extent to which Tregs can suppress the anti-tumor immune response in cancer patients. Implementing a larger sample size, CD24 antibodies, confocal microscopy, and flow cytometry in future studies would enable stronger conclusions to be drawn in regards to the effect of exercise on Treg populations in cancer patients. Further examination of the interactions between cancerous cells and various lymphocytes may greatly contribute to the development of more efficacious cancer treatment strategies in the future.
INTRODUCTION
In 2017, an estimated 1.7 million people were diagnosed with cancer, and approximately 600 thousand people died from it in the United States alone1. By 2030, an expected 21.7 million new cancer cases and 13 million cancer deaths will occur worldwide2. As the number of diagnoses and fatalities is only predicted to increase, further research on cancer is vital for the development and implementation of more effective methods of treatment.
Cancer is a disease characterized by the uncontrollable growth of cells, resulting in tumors. Malignant tumors in the body, caused by several mutations in DNA, are also capable of metastasizing to sites beyond the primary tumor. To further promote and sustain the growth of cancerous cells in tissue, tumor angiogenesis is induced: cancer cells influence their microenvironment by inducing nearby normal cells to form blood vessels so that the tumor’s access to oxygen and nutrients is maximized3.
In response, various white blood cells of the immune system, also known as lymphocytes, target and attempt to eliminate cancerous cells; however, through their interactions with specific lymphocytes, cancer cells can manipulate the immune system into defending the growth of a tumor and prevent the immune system from attacking these cells3.
Regulatory T cells (Tregs) are a subpopulation of T lymphocytes that modulate the immune system accordingly and suppress self-reactive lymphocytes to prevent autoimmune disease4. Treg cells work to maintain immune homeostasis so that other lymphocytes do not damage healthy tissue, yet this very process can aid the progression of cancer in a cancer patient. Though the primary function of Foxp3-expressing Treg cells is to inhibit immune response against self-antigens, in cancer patients, they have been found to subdue the anti-tumor immune response and induce tumor-specific local immune tolerance5.
Foxp3, the transcription factor responsible for the differentiation of Treg cells, is essential for the Treg suppressor function. Furthermore, in Treg precursor cells, Foxp3 stabilizes molecular features that affect cell function and maintenance and also reverses features that are detrimental to Treg cell function6. Therefore, the Foxp3 transcription factor is considered to be the most reliable marker for Tregs and consequently will be utilized as a biomarker to identify Tregs in this study7. CD25 is also a protein of interest in this study because it is a component of one of the cellular pathways that upregulates Foxp3. In this pathway, the signaling molecule IL-2 binds to the IL2RA receptor, otherwise known as the transmembrane protein CD25. This signaling activates the JAK protein, which subsequently stimulates the STAT5 protein. STAT5 increases Foxp38.
Tregs expressing the transcription factor FoxP3 are highly immune-suppressive, modulate tumor angiogenesis, and promote tumor progression by suppressing the activation and expansion of tumor-antigen-specific effector T cells. Because the presence of these regulatory T cells in the tumor microenvironment becomes protumorigenic, or advantageous for the growth of the tumor, cancer cells specifically recruit and direct these Tregs to impede antitumor immunity in the tumor microenvironment for defense against the body’s own immune system9. In fact, tumor-associated antigens may drive an early and rapid expansion of Tregs, since increased counts of CD4+CD25+ Treg cells have been observed in the tumor tissues of several cancers including lung, breast, ovarian, liver, pancreatic, and gastrointestinal10. As a result, high populations of Tregs in tumor tissues are frequently linked to poor prognoses in cancer patients.
However, there is growing evidence that the removal of Treg cells is able to evoke and enhance the anti-tumor immune response5. In a mouse model, systemic depletion of Treg cells 15 days after tumor implantation resulted in improved long-term survival. In a highly metastatic breast cancer model, lung metastasis was prevented through the depletion of Treg cells11. Because of this, depletion of suppression-competent Foxp3-high effector Treg cells is currently being tested extensively and even considered an effective cancer immunotherapy treatment strategy for some cancers12.
Interestingly, in addition to mitigating the adverse effects of invasive treatments such as chemotherapy, exercise has been shown to have a considerable impact on immune competence, lymphocyte populations, and lymphocyte proliferative response13. There is now evidence suggesting that exercise reduces the risk of primary cancer development, cancer recurrence, and cancer-specific mortality1415. Regular exercise has also been reported to regulate, deter, and in some cases even regress the growth of tumors16.
Prior research demonstrates that exercise has a beneficial impact on various aspects of cancer. However, definitive explanations regarding the mechanism by which exercise achieves these anti-tumor effects have yet to be identified. It is thought that exercise influences the proliferation of specific lymphocytes in the tumor microenvironment and lymphoid tissues, thereby influencing characteristics of the cancer. However, little research has been done to investigate exercise’s effect specifically on such lymphocytes. Consequently, this research project is concerned with the effect of exercise on regulatory T cell populations in the tumors and spleens of tumor-bearing rats. A decline in Treg population would limit the suppression of the anti-tumor response and in turn inhibit tumor growth; the latter result has been observed in cancer patients as an effect of regular exercise. Because of these findings, it is hypothesized that exercise will decrease the number of Treg cells present in the tumor and lymphoid tissue of tumor-bearing rats. The aim of this study, as hypothesized, is to observe decreased counts of regulatory T cells attributable to physical activity by utilizing Foxp3 and CD25 as biomarkers to identify Tregs in fluorescence microscopy images of rat tumor and spleen.
MATERIALS AND METHODS
Tumor Growth and Euthanization
Six rats were assigned to be either sedentary or exercising, with three rats in each cohort. Subjects were administered flank injections with MTLn3 rat mammary adenocarcinoma cell line 4 weeks after placement in cages. After injection, tumor growth was monitored and the voluntary wheel-running of exercising rats was recorded using cycle counters. While rat health and growth were observed regularly, rats were handled with care and given adequate amounts of food and water. Twenty-one days post-injection, or when tumors grew to 3 centimeters, rats were euthanized using sodium pentobarbital in an effort to minimize the burden of the tumor on the animal in a humane manner.
Dissection and Tissue Storage
Following euthanization, spleen and tumor tissue was excised from rats. Removed tissue samples were cut into small pieces, placed in cryomolds, coated in OCT compound, and frozen with isopentane and liquid nitrogen. Cryomolds were stored at
-80°C.
Cryosectioning
From storage, tissue samples were cryosectioned to 4 microns at 20°C. Cryosectioned tissue was placed onto 8 poly-D-lysine-coated slides. Each slide consisted of 2 tumor samples and 2 spleen samples. 1 tumor sample and 1 spleen sample on every slide was designated to be the negative control while the remaining 1 tumor sample and 1 spleen sample were designated to be stained. 2 slides were cut for each rat.
Immunohistochemistry
Tissue was fixed in acetone at -20°C in order to prevent the dissociation of the tissue. Tissue samples were then washed with TBS (0.05% Tween 20 in PBS) to help dissociate the cell membranes. Permeabilization of the membranes was accomplished utilizing a detergent comprised of Tween 20 and 0.1% Triton X-100. The blocking buffer, 1% NGS in PBS was applied to the tissue samples to increase antibody affinity. Positive control tissue samples were then stained with antibodies for CD25 and Foxp3 at a concentration of 1:200 for 1600 microliters. After being held at 4°C for 1 hour, the samples were washed in TBS. Hoechst dye was applied to all tissue samples for 10 minutes to stain the cell nuclei. Cover slips were fitted using 50% glycerol.
Visualization Using Fluorescence Microscopy
For each sample, 2 fields of view were randomly selected and observed in 40x magnification. Fluorescence microscopy was utilized to recognize CD25 and Foxp3 in order to identify Tregs and subsequently quantify the number of regulatory T cells present in each of the positive control tissue samples.
Fluorescence microscopes emit wavelengths of light that excite the fluorophores, marking particular antibodies. The absorbance of light by these fluorophores results in the emission of energy, which is then visualized through the use of a filter designed specifically for that wavelength. In this experiment, CD25 antibody fluoresced red under the PE filter, Foxp3 antibody fluoresced green under the FITC filter, and Hoechst dye fluoresced blue under the DAPI filter.
RESULTS

Figure 1. (Positive Control) Each tissue sample necessitated the consideration of all three fluorescence. In the fluorescence microscopy images (Figures 1, 2, 3, and 4), the upper right quadrant, upper left quadrant, and lower left quadrant display the presence of Foxp3, CD25, and cell nuclei, respectively. The lower right quadrant is an image overlaying the fluorescence of the CD25 antibody, Foxp3 antibody, and Hoechst dye applied. Points at which all three fluoresced are CD25+Foxp3 cells in phenotype and were identified as Tregs.
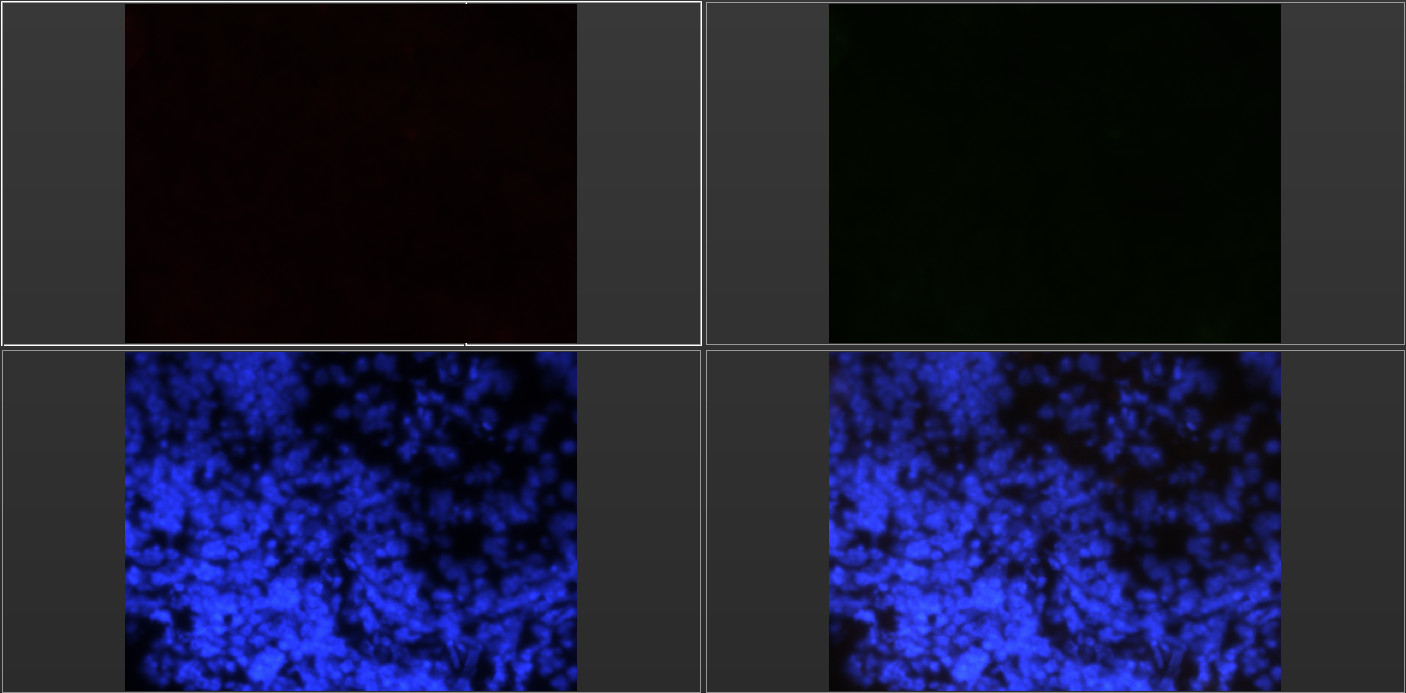
Figure 2. (Negative Control) Consists of fluorescence microscopy images of a negative control tissue sample. These samples were only stained with Hoechst dye. Consequently, there is no fluorescence of CD25 or Foxp3 in the upper two quadrants. Only cell nuclei fluoresced. Negative control samples were visualized to verify that the antibodies applied were responsible for causing CD25+ and Foxp3+ fluorescence.

Figure 3. (Positive Control – CD25+ Foxp3+ Cells) Contains fluorescence microscopy images of a positive control tissue sample. The green, red, and blue fluorescence is indicative of the presence of Foxp3, CD25, and nuclei, respectively. Cells such as those in the lower right quadrant that are boxed in red were identified as CD25+ Foxp3+ regulatory T cells.

Figure 4: (Positive Control – CD25- Foxp3+ Cells) Displays fluorescence for the same field of view presented in Figure 3. However, the cell boxed in the lower right image was identified as a CD25- Foxp3+ cell because it fluoresced green and blue but not red.
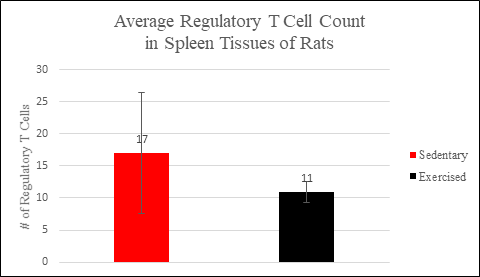
Graph 1. Compares the average regulatory T cell count in the spleen samples of the sedentary group to the average regulatory T cell count in the spleen samples of the exercised group. More regulatory T cells were present, on average, in the lymphoid tissue of the sedentary group than in the lymphoid tissue of the exercised group. Error bars represent one standard deviation above and below the calculated mean.

Graph 2. Compares the average regulatory T cell count in the tumor samples of the sedentary group to the average regulatory T cell count in the tumor samples of the exercised group. Considerably more regulatory T cells were present, on average, in the tumor microenvironment of the sedentary group than in the tumor microenvironment of the exercised group. Error bars represent one standard deviation above and below the calculated mean.

Graph 3. Compares the average regulatory T cell count in the tissue samples (spleen and tumor combined) of the sedentary group to the average regulatory T cell count in the tissue samples of the exercised group. Substantially more regulatory T cells were present, on average, in the tissues of the sedentary group than in the tissues of the exercised group. Error bars represent one standard deviation above and below the calculated mean.
DISCUSSION
The spleens of sedentary rats had on average 17 Tregs, with a standard deviation of 9, compared to the spleens of exercised rats which had on average 11 Tregs, with a standard deviation of 2. The tumors of sedentary rats had on average 16 Tregs, with a standard deviation of 14, whereas the tumors of exercised rats had on average 3 Tregs, with a standard deviation of 2. Overall, 33 Tregs were counted in sedentary rats as opposed to 14 Tregs in exercised rats.
Assessing the spleen samples and tumor samples of both cohorts, it was noted that sedentary rats had an increased amount of Tregs in their spleens and tumors compared to those of the exercised rats; however, the difference in Treg cell count between sedentary and exercised cohorts was considerably greater for the tumor tissue than for the spleen tissue. One potential implication of this observation is that exercise limits the recruitment of Tregs to the tumor microenvironment to a greater extent than it limits the mere presence of Tregs in lymphoid tissue. This is important because it is the population of Tregs in the tumor microenvironment that induces tumor-specific local immune tolerance, not merely the presence of excess Tregs circulating throughout the blood and lymphoid tissue; therefore, accumulation of Tregs in the tumor microenvironment is of greater concern and more relevant to the health and prognosis of a cancer patient. Moreover, the more constrained effect on Treg populations in lymphoid tissue may even be encouraging as it indicates that physical activity does not compromise beneficial Treg function, including the prevention of autoimmune disease, within a cancer patient. Though most of the data sets contained high standards of deviation, there was greater variability in the data concerning the tissues of sedentary rats, supporting the notion that exercise can help to reduce the unpredictability of Treg populations within cancer patients. Greater predictability can afford doctors greater control over the outcomes of the treatment process, and in turn, more success with the implementation of certain treatment strategies.
The lack of statistically different means, partly due to the high variability of Treg count in sedentary rats, can be attributed primarily to the limited sample size. Due to time constraints, tissue samples from only six rats could be collected and studied. Even so, this data indicates the existence of a relationship between exercise and smaller Treg populations in individuals with cancer: the tumor tissue of the exercised rats contained considerably fewer regulatory T cells than the tumor tissue of their sedentary counterparts.
Increasing the sample size would provide more points of data, thereby reducing the variability of the data amassed and enabling more descriptive statistics and stronger conclusions to be drawn. Additionally, working with the cancer line MATB3 would have been preferable to using the MTLn3 cancer line; while the MTLn3 cancer line is more aggressive, the development of the MATB3 cancer in the rats would have been more comparable to the progression of cancer in humans. Moreover, a novel Foxp3 immunohistochemsitry (IHC) protocol was designed and implemented for the rats. As a result, inherent flaws in the protocol were subject to adjustments and improvements throughout the course of this study.
A point of interest in this research was the identification of CD25- Foxp3+ cells. Although the transmembrane protein CD25 is present in many different lymphocytes, previous literature suggests CD25 to be a characteristic of Treg cells. However, these Foxp3-expressing cells did not fluoresce for CD25, indicating a relatively high population of another lymphocyte in the tumor microenvironment. The role of these cells in either the advancement or suppression of tumor growth should be considered for further study.
CONCLUSION
Overall, the data exhibited that the tissues of exercised rats contained, on average, decreased counts of Tregs as compared to the tissues of sedentary rats, suggesting that exercise regulates the quantity of Tregs and thereby limits the extent to which Tregs can suppress the anti-tumor immune response in cancer patients. Because prior literature determined that elevated levels of Tregs in patients correspond to poor prognoses, exercise may improve the prognoses of cancer patients by way of modulating the proliferation of Tregs and, consequently, moderating protumorigenic activity in the tumor microenvironment.
One substantial limitation of this trial was the small sample size, consisting of only 3 rats in each cohort. To extend this research, more trials should be conducted with a greater number of test subjects to obtain a more accurate understanding of the relationship between exercise, Tregs, and prognoses of cancer patients. For further replications of this research, applying CD24 antibodies to tissue samples would be valuable to help differentiate between Tregs and Bregs. Bregs are regulatory B cells that also participate in the suppression of immune responses. It would also be beneficial to use alternative forms of data collection in future studies. Confocal microscopy for IHC would provide clearer, more magnified views of the cells to more accurately locate CD25+ Foxp3+ cells; flow cytometry would enable greater amounts of quantitative data to be collected, allowing for more comprehensive and accurate analysis to be conducted.
In this experiment, an average of 33 Tregs were identified in the tissues of sedentary rats; however, the number of Tregs found in the tissues of exercised rats was 14, approximately 42% of the count for the sedentary cohort. The reduction of Tregs in the tumor microenvironment observed in this study can be considered and further explored as a mechanism by which exercise impacts cancer. More extensive research of this nature can be utilized to produce methods of treatment that mimic the tumor-regressive effects of exercise, significantly contributing to the development of more effective cancer treatment strategies and resulting in more favorable treatment outcomes. The implications of such research would improve prognoses for the millions of people diagnosed with cancer every year.
ACKNOWLEDGEMENTS
I am very grateful to the Frontiers of Science Institute at the University of Northern Colorado for providing me with the opportunity to engage in such interesting research, as well as Brian Davidson Health Sciences and Xcel Energy for sponsoring my research. I would like to thank Brian Krum for designing and mentoring my research, as well as Aspen Hirsch and Dr. James Haughian (at the University of Northern Colorado). I am also extremely appreciative of Shane Murphy at the University of Northern Colorado and Alana Stevenson at Cherokee Trail High School for reviewing and commenting on the contents of this study. Additionally, I am extremely thankful to Vivienne Lacy for being such an excellent research partner and supportive friend.
REFERENCES
-
Siegel, R. L., Miller, K. D., & Jemal, A. (2017, January 05). Cancer statistics, 2017. Retrieved July 02, 2017, from http://onlinelibrary.wiley.com/doi/10.3322/caac.21387/full
-
Global Cancer Facts & Figures. (n.d.). Retrieved July 2, 2017, from https://www.cancer.org/research/cancer-facts-statistics/global.html
-
National Cancer Institute. (n.d.). Retrieved July 02, 2017, from https://www.cancer.gov/about-cancer/understanding/what-is-cancer
-
Morris, S. Y. (2017, January 30). Everything You Should Know About Lymphocytes. Retrieved July 02, 2017, from http://www.healthline.com/health/lymphocytes#function2
-
Tanaka, A., & Sakaguchi, S. (2016, December 20). Regulatory T cells in cancer immunotherapy. Retrieved July 02, 2017, from http://www.nature.com/cr/journal/v27/n1/full/cr2016151a.html
-
Rudensky, A. Y. (2011, May). Regulatory T Cells and Foxp3. Retrieved July 2, 2017, from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3077798/
-
Corthay, A. (2009, October). How do Regulatory T Cells Work? Retrieved July 02, 2017, from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2784904/
-
Zorn, E., Nelson, E. A., Mohseni, M., Porcheray, F., Kim, H., Litsa, D., . . . Ritz, J. (2006, September 01). IL-2 regulates FOXP3 expression in human CD4 CD25 regulatory T cells through a STAT-dependent mechanism and induces the expansion of these cells in vivo. Retrieved July 2, 2017, from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1895505/
-
Wang, H. Y., & Wang, R. (2007, February 15). Regulatory T cells and cancer. Retrieved July 02, 2017, from http://www.sciencedirect.com/science/article/pii/S095279150700012X
-
Liu, C., Workman, C. J., & Vignali, D. A. (2016, March 01). Targeting regulatory T cells in tumors. Retrieved July 02, 2017, from http://onlinelibrary.wiley.com/doi/10.1111/febs.13656/full
-
Ha, T. (2009, December). The Role of Regulatory T Cells in Cancer. Retrieved July 02, 2017, from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2816955/
-
Fuerst, M. L. (n.d.). Targeting Regulatory T Cells for Cancer Immunotherapy : Oncology Times. Retrieved July 02, 2017, from http://journals.lww.com/oncology-times/fulltext/2016/11100/Targeting_Regulatory_T_Cells_for_Cancer.13.aspx
-
Rajarajeswaran, P., & Vishnupriya, R. (2009). Exercise in cancer. Retrieved July 02, 2017, from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2885882/
-
Brown, J. C., Winters-Stone, K., Lee, A., & Schmitz, K. H. (2012, October). Cancer, Physical Activity, and Exercise. Retrieved July 02, 2017, from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4122430/
-
Betof, A. S., Dewhirst, M. W., & Jones, L. W. (2013, March). Effects and potential mechanisms of exercise training on cancer progression: a translational perspective. Retrieved July 02, 2017, from https://www.ncbi.nlm.nih.gov/pubmed/22610066
-
Hoffman, S. A., Paschkis, K. E., DeBias, D. A., Cantarow, A., & Williams, T. L. (1962, June 01). The Influence of Exercise on the Growth of Transplanted Rat Tumors. Retrieved July 02, 2017, from http://cancerres.aacrjournals.org/content/22/5_Part_1/597.short
- Siegal, R. L.; Miller, K.D.; Jemal, A. Cancer statistics, 2017. 2017. [↩]
- American Cancer Society [↩]
- National Cancer Institute [↩] [↩]
- Morris, S. Y. Everything You Should Know About Lymphocytes. 2017. [↩]
- Tanaka, A.; Sakaguchi, S. Regulatory T cells in cancer immunotherapy. 2016. [↩] [↩]
- Rudensky, A. Y. Regulatory T Cells and Foxp3. 2011. [↩]
- Corthay, A. How do Regulatory T Cells Work?. 2009. [↩]
- Zorn, E.; Nelson, E. A.; Mohseni, M.; Porcheray, F.; Kim, H.; Litsa, D.; Bellucci, R; Raderschall, E.; Canning, C.; Soiffer, R.; Frank, D.; Ritz, J. IL-2 regulates FOXP3 expression in human CD4 CD25 regulatory T cells through a STAT-dependent mechanism and induces the expansion of these cells in vivo. 2006. [↩]
- Wang, H. Y.; Wang, R. Regulatory T cells and cancer. 2007. [↩]
- Liu, C.; Workman, C. J.; Vignali, D. A. Targeting regulatory T cells in tumors. 2016. [↩]
- Ha, T. The Role of Regulatory T Cells in Cancer. 2009. [↩]
- Fuerst, M. L. Targeting Regulatory T Cells for Cancer Immunotherapy : Oncology Times. [↩]
- Rajarajeswaran, P.; Vishnupriya, R. Exercise in cancer. 2009. [↩]
- Brown, J. C.; Winters-Stone, K.; Lee, A.; Schmitz, K. H. Cancer, Physical Activity, and Exercise. 2012. [↩]
- Betof, A. S.; Dewhirst, M. W.; Jones, L. W. Effects and potential mechanisms of exercise training on cancer progression: a translational perspective. 2013. [↩]
- Hoffman, S. A.; Paschkis, K. E.; DeBias, D. A.; Cantarow, A.; Williams, T. L. The Influence of Exercise on the Growth of Transplanted Rat Tumors. 1962. [↩]



{kind=link}
{kind=link}