
Abstract
The aspect ratio, a measure of “wing pointedness,” is a fundamental measure of wing shape. Though it has been extensively studied in individual fields, few studies have directly compared its effects across both engineering and biology. This paper aims to address this gap by examining how AR influences flight performance in both biological and mechanical flight, with an emphasis on its application to morphing-wing multi-mission vehicles. A systematic literature review was conducted, consisting of peer reviewed papers sourced from Google Scholar, Science Direct, and other databases. The findings suggest that high ARs correlate to flight efficiency, while low ARs improve maneuverability, with similar patterns observed across biological and mechanical flight systems. However, studies on insect flight are less conclusive. Possible models and designs for morphing-wing multi-mission vehicles are also considered. Finally, current mechanical designs with morphing wings are considered, and found to fail at simultaneously morphing the AR and other critical airfoil parameters
Introduction
The AR, a ratio between the wing chord and span, has long been considered a fundamental parameter of wing shape. Extensive research into the AR already exists in planes, birds, and insects, although the underlying mechanics of its effects on flight still are not entirely understood1,2,3,4,5,6. The large majority of this research is restricted to a single discipline, either engineering or biology. In the engineering field, many papers consider only rigid, non-flapping wings, like those found on most conventional planes. Literature focused on biological organisms, like birds and insects, include a much broader range of flight styles, including gliding, flapping, and diving. In recent years, interest in biologically inspired flight vehicles has increased, leading to more interdisciplinary papers considering biological flight mechanics for planes and other flight vehicles. Some of these papers have focused on the wing morphing capabilities of birds and insects, considering how these mechanisms could be adapted to plane or drone flight.
Despite increased interdisciplinary research, few studies directly compare AR effects in engineering and biology. To support future flight vehicle designs with either flapping and rigid wings, this paper seeks to conduct an interdisciplinary review on research into the AR. Compiling AR research gives designers of future flight vehicles confidence in the function of the AR in both rigid and flapping flight. In addition, considering birds and insects shows the implications of different ARs in real world environments, rather than the ideal conditions assumed in aerodynamic equations. This paper then considers the AR morphing abilities of birds and insects to evaluate what biological mechanisms could be adapted to actively change the AR in mechanical flight. Actively adapting the AR according to environmental conditions could allow flight vehicles to reap the benefits of both low and high ARs. Finally, this paper looks at current progress in AR morphing designs, and considers what future designs must incorporate to further increase flight capabilities.
Though the AR is an important parameter of wing shape, it does not consider many important determinants of flight performance, like 3D wing configuration, camber, twist, mass, and many others. Flight performance in birds is heavily impacted by their tails, which can change shape and create aerodynamic forces not measured by the AR. Therefore, the correlations between flight performance and the AR researched in this paper may be impacted by other factors as well, and wings of a certain AR are not guaranteed to perform as predicted. In addition, this paper does not consider the changes in AR while flapping. By flapping, birds radically retract and extend their wings, significantly changing their effective AR. However, these AR changes are simply a byproduct of lift generation, rather than a reaction to environmental conditions. Instead, this paper focuses on gliding and diving flight, in which the AR is morphed specifically in reaction to environmental changes, or to perform a particular maneuver. Therefore, this paper does not consider ‘short term’ changes, restricting the kinds of wing morphing included.
This paper will first give an overview of basic flight mechanisms, biological flight, and multi-mission flight vehicles. Then, it will review current research on the AR in planes, birds, and insects. From there, it will consider the AR morphing capabilities of birds and insects, before outlining current progress in mechanical wings with morphing ARs.
Materials and Methods
This article conducts a systematic literature review of the aerodynamic effects of the aspect ratio (AR) in both biological and engineering contexts, as well as the morphing capabilities of AR in birds, insects, and airplanes. The review includes studies that address the effects of AR on flight dynamics, biological wing morphing, and technologies related to wing morphing in engineered systems.
Articles were found through searches in Google Scholar, Science Direct, and Google, using both individual and combined keywords. Each database was last consulted on December 5th, 2024. Terms used to identify AR-related studies were ‘aspect ratio,’ ‘high/low aspect ratio,’ ‘hand wing index,’ ‘wing shape,’ ‘wing span,’ ‘wing area,’ ‘bird flight,’ and ‘insect flight.’ Terms used to find studies on wings with morphing ARs were ‘wing morphing,’ ‘flexibility,’ ‘changing wingspan/wing area,’ and ‘morphing aircraft/vehicles.’ Boolean operators (AND, OR) were applied informally where possible, depending on the search platform.
The search was restricted to journal articles, textbooks, book chapters, and patents on mechanical wing-morphing designs. No language restrictions were applied, but articles published between 2000 and 2024 were given preference, particularly for wing morphing research, to reflect recent developments in the field.
Articles were first screened for relevance in the title and abstract, before the full text was assessed. Studies were included if they provided detailed information on the aerodynamic effects of AR or described a biological or mechanical wing morphing mechanism.
All reports were screened and reviewed by a single reviewer, and no automation tools were used at any stage of the search, screening, or data extraction process. No formal risk of bias assessment or quality appraisal of included studies was carried out, which is acknowledged as a limitation of this review.
Results
Theory of Flight
Flight has been a part of the natural world for hundreds of millions of years. Generally, flight performance is constrained by four main forces: lift, drag, thrust, and weight.
Lift is a force in the upwards direction, used to overcome the force of gravity. In most airfoils, lift is generated through the flow of air over a wing. The shape of the airfoil speeds up the air above it, creating a pressure difference above and below the wing. Because particles in high-pressure areas tend to move towards low-pressure areas, this imbalance pressure distribution creates a net force in the lift direction. One explanation for this pressure imbalance is Newton’s Third Law, which states that for every action, there is an equal and opposite reaction. As the air flows over an airfoil, the airfoil displaces large amounts of air, creating an upwash and a downwash. To create the downwash, the airfoil must be exerting a downwards force on the air. According to Newton’s Third Law, this means the air exerts an upwards force on the airfoil in response, generating lift. The pressure imbalance can also be explained through Bernoulli’s Principle and the continuity equation.
The continuity equation:
(1) 
(2) 
where p is pressure, V is velocity,
and A is the cross-sectional area of flow.
Bernoulli’s Principle:
(3) 
(4) 
where p is static pressure of the fluid at the cross section, p is density of the flowing fluid, g is acceleration due to gravity, v is mean velocity of fluid flow at the cross section, h is the elevation head of center at the cross section.
Most airfoils are asymmetrical, meaning more air is displaced above the airfoil than below. According to the continuity equation, this displacement or loss of flow area means an increase in velocity. Bernoulli’s Principle is another version of conservation of energy, stating that the sum of the pressure energy, kinetic energy, and potential energy is always constant. According to this equation, an increase in velocity equals a decrease in pressure. Therefore, the air displaced by the airfoil leads to an increase in velocity, and a decrease in pressure, generating the pressure distribution and lift. Although airfoils can be specialized to generate more lift, any object that creates a pressure distribution will generate lift. However, there are other forces working on the airfoil as well.
Drag is any force opposite to the direction of an object’s movement, and comes from many different sources. In flying objects, the drag is mainly comprised of parasite drag and induced drag. Parasite drag stems from the shape and surface area of the airfoil. Skin friction, or the friction caused by the flow of air particles over an airfoil, is one kind of parasite drag. Parasite drag generally depends on the shape and surface area of an airfoil, as well as the velocity and density of the air. Similarly, profile drag refers to the drag created by the shape and surface of the wing. Induced drag is an inevitable byproduct of lift, and typically stems from vortices and downwash. The total aerodynamic drag on an airfoil is the sum of the parasite, profile and induced drags.
Weight is the downward force on an object due to gravity. The heavier a plane or organism, the more lift is required to keep it off the ground. Therefore, plane design is often a struggle between aerodynamically ideal configurations and weight. Biological organisms are evolved to reduce their weight as much as possible, while still retaining flight capabilities. Birds, for example, have bones with tiny air pockets that help reduce their weight.
Thrust is the force that pushes an object forward through the air. In planes and rockets, thrust is mainly generated through the combustion of fuel, while thrust in organisms is generated through the flapping of wings. Thrust is essential to maintaining lift, which depends on the flow of air over the wing.
Key Components of Airfoils
Airfoils are shaped by several different components: the leading/trailing edge, the angle of attack, the chord, the camber, and others. The leading and trailing edges, which are the front and back of an airfoil respectively, affect the upwash and downwash generated, and also assist in the creation of vortices. The chord is the straight line between the leading and trailing edge. The camber is the curve of an airfoil’s top and bottom surfaces.
An airfoil’s angle of attack (AOA) is the angle at which it meets the incoming airflow. A steeper angle of attack increases both the lift and the drag of an airfoil. At slow speeds, flight vehicles and organisms will often use high AOAs to generate enough lift to remain airborne, even with decreased airflow. Eventually, if an AOA becomes too steep, it can cause flow separation. Flow separation is when the air particles along the bottom of an airfoil detach, moving the high- pressure air further away, and reducing lift. To counteract flow separation, some airfoils generate vortices that help fill this gap, and drag high pressure air back towards the low-pressure air.
Vortices, or rotating masses of air, are a byproduct of lift. The pressure distribution created by the airfoil causes high pressure air from below to curl over the edge of the wing, into the low- pressure area. This curl creates a vortex off the edge of the wing (Fig. 1). Other parts of airfoils can also be designed to create vortices. Vortices primarily produce drag, specifically induced drag, due to the turbulence generated by the swirling air. However, vortices can also indirectly generate lift, especially in wings with high-lift, like those on birds and insects. Leading edge vortices (LEVs), for example, can delay flow separation at high AOA, ensuring continued lift generation. Under some circumstances, LEVs can also help create low-pressure regions on top of the wing, increasing the lift generated.

Flight Efficiency
Flight efficiency refers to the ability of an aircraft to maximize its performance, while minimizing expended energy. Performance may refer to distance traveled, speed achieved, maneuverability, or a myriad of other applications for flight.
Flight efficiency is most often measured through the lift to drag (L/D) ratio, or glide ratio. A higher L/D ratio means the flight vehicle generates more lift relative to drag, and therefore needs less input energy to sustain forward flight. This means that designers looking to optimize for flight efficiency will either look to increase lift, or decrease drag.
The lift and drag coefficients are highly related to flight efficiency. The lift coefficient is a dimensionless number that quantifies the efficiency of a wing in generating lift. It is affected by the lift force, air density, velocity, and wing area. Similarly, the drag coefficient is a dimensionless number measuring the drag experienced by an airfoil, calculated from the drag force, air density, velocity, and wing area. These coefficients are highly related to the L/D ratio, and can be used to quantify flight efficiency. A large lift coefficient means an airfoil efficiently generates a large amount of lift, and likely has a high L/D ratio. A low drag coefficient means an airfoil generates very little drag under specific conditions, also increasing the L/D ratio.
Biomechanical Flight
Many planes, drones, and other machines designed for flight have been created through imitating biological processes. The aerodynamic and mechanical complexities of biomechanical flight have made it extremely hard to recreate exactly. Like man-made wings, biomechanical wings create pressure distributions to generate lift. However, biological organisms cannot use propellers or engines to generate thrust. Instead, birds and insects flap their wings to generate both lift and thrust. Biological flight consists of a mix between flapping and gliding, often actively alternating between the two.
Bird Flight
Birds generate powerful aerodynamic forces to keep themselves aloft via flapping. This movement is primarily powered through strong muscles in the bird’s breast, which contract and extend to perform each flap. On the downstroke, bird wings deflect air downwards and behind them, generating upwards and forwards forces according to Newton’s 3rd Law; by maintaining a small angle between their wingstroke and flight direction, they generate both thrust and lift on the same stroke, conserving energy and minimizing their wing-stroke frequency8. On the upstroke, birds hinge their wings and separate their feathers, allowing air to pass through more easily, reducing drag. Though most birds stay within a set range of wing-beat frequencies, some can alter theirs at will5.

Like airplanes, birds adjust their energy expenditure to maintain different flight speeds. At low speeds, reduced lift forces them to increase their wingbeat frequency, thereby using more energy5. As their speed builds, the wings generate more lift, allowing the birds to decrease their wingbeat frequency and conserve energy. However, at very high speeds, birds must once again increase their wingbeat frequency to produce more thrust, leading to higher energy use. As a result, their power curve typically forms a U-shape, similar to that of airplanes, although some species deviate from this pattern in Fig. 2. For migratory birds, the most energy-efficient speeds often align closely with their migration velocity, as their limited fuel reserves demand maximum efficiency.
Birds must twist and alter the shape of their wings to retain an optimal configuration while flapping. For example, they must twist their wings to flap while also engaging optimal angles of attack and incidence10. This twisting is made possible through the bird wing’s flexible joints and feathers. The structure of bird wings resembles that of a human arm and hand, including flexible joints that allow the wing to twist. Finger-like digits throughout the wing also allow birds to actively change the shape of their wings. The wing feathers are attached to the bones through ligaments, and form the wing surfaces. Feathers are layered across the wing, capable of sliding beneath each other and retracting. As the ‘arm’ is extended, the feathers are automatically spread, allowing the bird to alter its wing shape. The feathers themselves are somewhat flexible, which contributes to wing twisting in flight11.
Constant flapping generates a lot of force but also takes large amounts of energy. In birds, only about 25% of metabolic energy is converted into mechanical energy, making flight efficiency extremely important5. To conserve energy, birds will either use their forward momentum to glide, or utilize updrafts to soar. To soar, birds either flap until they reach a certain altitude and speed, or jump from a high perch. Then, they are pulled down by gravity, leading to airflow over their wings and thus lift. In stagnant air, gliding leads to a slow descent for the bird, eventually forcing the bird to land and restart. However, in areas where the force of updrafts equals the force of gravity, birds can continue soaring indefinitely, without expending excess energy. Birds can catch a wide range of updrafts, natural and man-made. Some utilize slope winds to cross mountainous regions; others capitalize on man-made winds, by flying near ferries or other vehicles; and some use thermals, rising columns of air generated by temperature differences due to the sun. However, these updrafts are highly inconsistent, forcing birds to regain altitude through flapping5.
Unlike planes, which are often rigid and unadaptable, birds actively morph their wings, allowing them to perform complex flight maneuvers and maintain high flight efficiencies. Through the flexibility of their joints and wing mobility, birds can change the camber and thickness of their wings, allowing them to actively adapt to environmental conditions10. Therefore, they are less affected by unsteady airflow, and can recover quickly from rapid maneuvers without succumbing to gravity or drag. Even when not actively controlled, flexible airfoils have been shown to create a thrust benefit of up to 50%12. In recent years, the scientific community has focused on attempting to emulate the adaptability of bird wings in mechanical design.
Insect Flight
Insect wings consist of a light, thin, and flexible membrane, supported by a series of veins. These veins function somewhat like a skeleton, maintaining the wing’s shape and distributing aerodynamic forces. The wings attach to the body through specialized, highly flexible joints, allowing for greater changes in configuration. Because they are made of one membrane, changes in insect wing shape are mainly due to twisting and flexing. However, some insects have more than 2 wings, allowing for some changes in wing area by adapting the overlap between the wings. Multiple wings also create higher surface area and stability, as well as increased maneuverability by moving all wings independently. Under some circumstances, having multiple wing pairs leads to increased flight efficiency, through higher lift generation and decreased turbulence.
Insect flight, like bird flight, consists of both flapping and gliding. However, only larger insects, that reach higher speeds and have larger wing areas, can properly glide. Most insects fly at slower speeds than birds, and are forced to constantly flap. Some research indicates that according to standard aerodynamic models, relying on steady-state equations for rigid airfoils, insects don’t generate enough lift to remain airborne13. The leading edge vortices (LEV) helps prevent airflow separation, delaying stall and indirectly increasing lift. LEVs may also help displace air downwards through their rotation, aiding in lift generation13.
Multi-mission Flight Vehicles
In recent years, interest in multi-mission flight vehicles has increased dramatically. Traditional planes, or drones, are optimized to perform one task at a time. Some aircrafts are designed for speed and maneuverability, while others are designed for long distance flight. Multi-mission flight vehicles would be designed to perform two missions in one flight, requiring a morphing of their flight configurations. ‘Hunter-killer’ aircrafts, meant to first find and then attack targets, are a classic example of multi-mission flight. These flight vehicles have many applications, including surveillance, environmental mapping, and military uses. Multi-mission vehicles cover a wide range of sizes, with special interest in micro air vehicles (MAVs), and nano aerial vehicles (NAVs)13. Small vehicles are compact, and able to access environments that larger vehicles cannot. However, due to their small size and mass, MAVs have little to no space for transmission devices, meaning they are often completely autonomous. In addition, MAV flight needs to be very efficient, as they do not have space for large energy stores. It is critical they can adapt to environmental conditions to preserve flight efficiency on their own, as birds and insects do.
Designers have turned to biomimetics—imitating natural flight mechanisms—to achieve morphing flight. Birds and insects adjust their wing shapes based on environmental conditions and aerodynamic needs. Falcons, for example, can both glide and execute steep dives, serving as a natural “hunter-killer” model. Since the size and weight of birds and insects are comparable to those targeted for MAVs and NAVs, their flight strategies are especially relevant. Many engineers aspire to replicate flapping capabilities found in nature, but doing so requires complex systems that incorporate morphing configurations, flexible materials, and adaptive control methods. Smart materials, such as shape-memory alloys and flexible composites, are crucial because they allow aircraft wings to change shape in real time. Although considerable research on morphing vehicles already exists, additional studies of biological flight strategies and advanced smart materials will be necessary to fully emulate bird- or insect-like flight.
Aspect Ratio
The AR compares the wingspan to the total wing area, and helps quantify the ‘pointedness’ of a wing. The AR is commonly used in both mechanical and biomechanical flight, and is closely related to the HWI in birds. It is one of the most important parameters of wing morphology, and is closely linked to the lift, drag, and efficiency of a wing.
(5) 
Where AR is the aspect ratio, b is the wingspan, S is the total wing area including the section of the body between the wings, and c is the mean wing chord
(6) 
Where  , i is induced drag,
, i is induced drag,  is the lift coefficient, and AR is the aspect ratio
is the lift coefficient, and AR is the aspect ratio
According to the equation (5), the wing area and wingspan have huge impacts on the AR. The wing area itself is impact by the wing chord, which therefore also influences the AR. If the wing area remains constant, and the wingspan increases, the AR will increase as well. However, if the wing area changes as well, the AR may increase or decrease, depending on how the extra span and area are added.
Subsonic Airplanes
According to the referenced equation, induced drag is inversely proportional to AR1. In other words, as the AR increases, induced drag decreases, which in turn raises the lift-to-drag (L/D) ratio. Consequently, conventional aircraft designs strive to increase AR to reduce induced drag. This relationship can be explained by the more favorable lift distribution seen in high-AR wings. Induced drag partly arises from the vortices formed at the wingtips. These vortices interact with the surrounding airflow, creating turbulence and contributing to drag. Because high-AR wings are longer, lift is spread more evenly across the span, resulting in smaller wingtip forces at each specific point and therefore weaker vortices. Smaller vortices mean less turbulence and induced drag, improving the wing’s overall efficiency14. For the same reason, favorable lift distribution, increasing the wingspan of a wing reduced induced drag as well. Moreover, a significant portion of lift originates at the leading edge of the airfoil. A wing with a higher AR has a greater wingspan, effectively increasing its leading edge length and thereby enhancing its total lift-generating capability1.
High AR wings have numerous applications. For instance, aircraft operating at high altitudes often employ higher AR wings to counteract the reduction in lift. Because gravity concentrates most of the air mass near Earth’s surface, air pressure drops dramatically at high elevations. Low air pressure reduces the number air particles flowing across the airfoil, weakening the pressure distribution and decreasing available lift. Airplanes at high altitudes must improve their flight efficiency to support their weight and remain airborne. As a result, they often feature high AR wings to decrease induced drag by shrinking the wingtip vortices, and increase the lift-to-drag (L/D) ratio or flight efficiency, thus offsetting the reduced air pressure and lift.
Despite these aerodynamic advantages, there is a practical limit to increasing a wing’s AR. As the AR grows, the wing becomes longer, thinner, and therefore more fragile. Beyond a certain point, the wing becomes too thin to support the lift forces, which may cause it to break. Increasing the AR also increases the bending moment at the wing root by moving the lift distribution farther from the main body. The lift forces on a wing pull it up and away from the main body, acting as a lever on the wing root. As the AR increases, these lift forces move further away from the wing root, thereby increasing the stress and the bending moment at the wing root. Consequently, increasing the AR requires a stronger wing root structure to withstand the stress. However, stronger structures are heavier and reduce the aircraft’s speed and efficiency. Due to the balance between aerodynamic design and structural integrity, most conventional subsonic aircraft have ARs ranging from 6 to 8. Within this range, the aircraft are fairly efficient while still maintaining the structural integrity needed for a reasonable degree of speed. Smaller ARs are also typically more practical for attaching fuel tanks, landing gear, and for storage in hangars. In contrast, some sailplanes designed for long-range flight have ARs in the 40s and 50s to minimize induced drag. Lower drag translates into reduced fuel requirements and weight, thereby improving efficiency1. Gliders like these typically have wings designed to distribute stresses and bend rather than break.
Although a high AR improves wing efficiency, it also diminishes maneuverability by giving the aircraft a high moment of inertia. The moment of inertia—resistance to changes in rotational speed—depends on the distance between the force and the center of rotation. In high AR wings, lift forces act farther from the aircraft’s centerline, increasing rotational inertia and centripetal forces, which lower the roll rate and agility1. In addition, having longer wings distributes an aircraft’s mass further from the center, increasing inertia and resistance to rolling or turning maneuvers.
The slender structure of high AR wings also makes them structurally suboptimal for complex maneuvers. Long, thin wings are more prone to twisting and flexing under sudden direction changes, reducing control and creating additional drag. While strengthening the wings themselves could stop bending, doing so would add mass and weight, requiring more force to execute turns and remain airborne, decreasing overall efficiency.
Aircraft that prioritize maneuverability often have low AR wings, which are shorter and stubbier. Low AR wings generate more induced drag, making them less suitable for long-range flight, but they offer increased agility due to their smaller pressure distribution. Because low AR wings are typically short and stubby, they have more spanwise flow than high AR wings, and more air can flow around the edge of the wing across the pressure distribution. This flow creates larger wingtip vortices, which increase drag but also contribute to a more gradual stall. Since low AR wings are shorter and rounder, they experience more spanwise airflow, which affects their pressure distribution. While this reduces overall lift, it helps them maintain lift at high angles of attack (AOA). As AOA increases beyond a certain point, airflow over the wing becomes disrupted, leading to stall. Low AR wings tend to experience early stall at the wingtips, but other sections of the wing often continue generating lift, resulting in a more gradual loss of lift rather than a sudden drop. In contrast, high AR wings have less spanwise airflow and a more even pressure distribution, which makes their stall behavior more abrupt. When they reach their critical AOA, they tend to stall uniformly across the wing, causing a sudden and more severe loss of lift. Because of this, low AR wings generally have a higher stall AOA than high AR wings15.
At slower speeds, reduced airflow requires increasing the AOA to maintain lift. Increasing the AOA deflects more air downwards, generating more lift, but makes flow separation and stall more likely. Low AR wings’ gradual stall makes them capable of generating lift at higher AOAs, making them more efficient for takeoffs, landings, and sharp turns. These maneuvers require the aircraft to slow down rapidly without losing lift, requiring a high AOA that doesn’t induce stall. Steep climbs benefit especially from high AOA and increased lift generation, as the entire maneuver consists of lift, making low AR wings especially valuable. Additionally, because stall occurs more gradually in low AR wings, pilots have more time to react and recover before losing control. While low AR wings incur more induced drag, they stall less easily at low speeds and are ideal for gliding flight. Their compact shape is also less prone to flexing and flutter, generally reducing structural weight compared to high AR designs2.
Wings with high wing areas typically stall at lower speeds, because they can generate more lift. As speed decreases, less air flows across the airfoil, decreasing lift. However, since wings with high wing areas can deflect more air, they generate more lift, and can sustain flight at slow speeds, decreasing a plane’s stall speed. Usually, low AR wings have higher wing areas, further helping them avoid flight at low stall speeds. While the wing area itself doesn’t directly affect induced drag, it does indirectly affect drag by playing a role in a wing’s AR. A low AR has stronger wingtip vortices, leading to more induced drag, while a high AR generates less drag.
In Birds
AR has been extensively used to examine bird flight. However, determining a bird’s wing area can be challenging, making AR measurements less widely available than other parameters. Additionally, AR can only be measured in live birds, not museum specimens, further limiting data availability.
Research on AR in birds generally supports the findings from mechanical flight studies. Many studies have linked high-AR wings to enhanced flight efficiency, particularly in migratory species3,4. Migratory birds, which routinely cover long distances, often have high-AR wings. According to Lockwood et al. and others, birds that depend on frequent or long-duration flights may have evolved high-AR wings to maximize efficiency6. Other research shows that AR is one of the best predictors of a bird’s natal dispersal distance—how far it travels from its birthplace to its first breeding site3. Because migratory birds typically have longer dispersal distances, this finding further connects high AR to flight efficiency. Although many studies focusing on AR and migration have examined passerines, investigations of other groups, such as shorebirds, have yielded similar results, suggesting a widespread trend16.
Some studies have begun linking high-AR wings to underlying aerodynamic principles. In birds, high-AR wings are associated with smaller wingtip vortices and reduced induced drag6. Modeling bird-like flapping has shown that high-AR wings can carry more weight over longer distances than low-AR wings, indicating greater efficiency17. In hummingbirds, increasing wing AR reduces the power required to hover18.
Because they are small and lightweight, birds can exploit air currents, such as thermals, slope winds, and even human-made updrafts, to dramatically reduce their power consumption while staying aloft. Many marine birds, which rely on wave and dynamic soaring, have evolved high-AR wings ideally suited for harnessing certain updrafts, thereby improving efficiency. Terrestrial thermal-soaring birds tend to have long, broad wings rather than extremely high ARs, but these differences may reflect habitat preferences rather than contradicting the link between soaring ability and high AR19. Even these broader-winged species often have a moderately high AR, with values around seven5.
Similar to airplanes, high-AR wings in birds maintain their efficiency only up to a point. For hummingbirds, once the AR surpasses about four, the lift-to-drag ratio declines again, reducing overall efficiency20. Researchers suggest that beyond a certain elongation, the wingtips become too pointed, making the wing more vulnerable to aerodynamic stress—paralleling the structural challenges of very high AR wings in airplanes6.
For small birds, maneuverability and agility are crucial, favoring the development of low-AR wings. Such wings have been linked to increased turning performance and lower inertia, aiding in rapid escapes or takeoffs6,21. Some birds use quick takeoffs and agile flight to evade predators or perform mating displays, leading to the evolution of shorter, more rounded wings. These rounded, low-AR wings generate a significant portion of their lift near the wingtips, where both the wing and air particles move faster. In species like the European starling (Sturnus vulgaris), having rounder wings significantly increases the angle of ascent, demonstrating the additional thrust and maneuverability provided by shorter wings6.
In Insects
As with studies in birds, AR is frequently used to quantify insect wing morphology. However, because insect wings vary greatly in shape, researchers usually rely on total wing area rather than mean wing chord to calculate AR. Unlike birds, many insects have four wings instead of two, changing how AR influences their overall wing planform and flight dynamics. For example, butterflies have four wings of relatively high AR; combined, these create a large total wing area relative to their body size. Generally, insects have lower ARs than birds because their wingspan is constrained by their body length. Insects typically exhibit ARs ranging from 1.5 to 6, whereas birds span a much wider range, from about 5.5 to 1922.
In insects, the relationship between high AR and lift efficiency is still under debate. Recent research has focused on the roles of the leading edge vortex (LEV) and trailing edge vortex (TEV), both of which influence the lift and drag produced by a wing. The LEV is a key mechanism insects use to generate significant lift. Its stable attachment is especially important: a larger, well-attached LEV creates a region of lower pressure over the wing’s upper surface, thereby increasing overall lift22. According to the Navier-Stokes equations, centripetal and Coriolis forces in flapping wings scale with AR and help maintain stable vortices. In insects, these forces stabilize the LEV during the wing’s sweeping motion (its angular displacement while flapping). Consequently, changes in AR affect the LEV and thus influence the wing’s lift and overall efficiency.
As AR increases, the LEV tends to become more compact, stable, and intense. This compaction increases the suction force (reducing pressure), enhances lift, and makes lift generation more uniform across the wing surface, ultimately reducing drag and improving flight efficiency. Correspondingly, experiments have shown that increasing AR raises the mean lift coefficient over a flap cycle23,24 and reduces the drag coefficient, resulting in a higher lift-to-drag (L/D) ratio24.
However, the benefits of increasing AR have limits. At an AR of about 1, the LEV forms a single stable vortex that compacts as AR rises. By AR 4, the LEV splits into dual vortices with a helical structure, indicating it is nearing a breakdown22,25. At even higher ARs, the LEV forms bubble-like structures that eventually burst into smaller vortices, reducing lift. This LEV breakdown may occur because high AR reduces centripetal and Coriolis forces, which are crucial for sustaining stable vortices and suction. Moreover, at higher AR, the wing chord is smaller, providing less surface area for the LEV. As a result, the LEV moves toward the trailing edge, causing vorticity from beneath the wing to evolve into a TEV, eventually shedding and producing induced drag25,26. Other studies note lift reductions at AR values of about 6, with a sharp decline at around 7.5, although such discrepancies may stem from testing different wing planforms26.
Some research has shown that lift can decrease entirely as AR increases. Studies directly comparing AR and lift coefficients have identified optimal efficiencies across a range of ARs, while others have found that interactions between the LEV and underside vortices reduce lift at higher AR values25. Local LEV detachment—where flow reverses at the trailing edge to form a TEV—occurs more rapidly at high AR. For instance, while an AR of 1.5 exhibits instability but no LEV detachment, higher ARs between 3 and 7 show progressively earlier detachment26.These inconsistencies may result from variations in how lift coefficients are calculated, differences in Reynolds numbers, and changes in Rossby numbers, rather than an intrinsic contradiction in the relationship between AR and lift22.
Hand-Wing Index
The Hand-Wing Index (HWI) quantifies the elongation of a wing, and relates Kipp’s distance (DK) to the length of the longest primary feather (LW) (Fig. 3). This index is very similar to the AR, although it focuses more specifically on the elongation of the wing hand. The HWI is less correlated to flight efficiency than other parameters, possibly because it ignores arm length, leading it to underestimate the elongation of birds with particularly long arms27. Nonetheless, the ease of calculating the HWI and its wide availability makes it a relevant parameter of wing morphology. Unlike the AR, the HWI doesn’t require the surface area of a wing, instead measuring specific distances. The HWI can be also measured from both avian museum specimens and live birds, making it especially useful for gathering more complete sets of data28. In addition, the measurements needed for the HWI are easier to measure reliably, likely making HWI data more accurate29.

(7) 
where  is Kipp’s distance, and
is Kipp’s distance, and  is the length of the longest primary feather.
is the length of the longest primary feather.
(8) 
The HWI and the AR of a wing are strongly correlated, as they both pertain to how elongated a wing is. One study comparing the HWI and AR of various Furnariidae species (Fig. 4) approximated an equation showing a positive linear relationship, indicating that an increase in the HWI meant an increase in the AR29. The AR can be related to the HWI through the equation:
(9) 
Though this equation is specific to furnariids, it indicates a strong linear correlation between the HWI and the AR, and allows us to use them as proxies for each other (Fig. 4). Therefore, the aerodynamic effects of the HWI apply to the AR as well, and vice versa.

Like the AR, the HWI has been correlated to flight efficiency, and used as an accessible proxy for efficiency in long-distance bird flight. A high HWI is correlated to a high natal dispersion, indicating that wings with a high HWI are more efficient30,31,32. Often, birds with a high HWI are migratory birds that travel long distances in search for food, mates, and other resources. However, a high HWI isn’t necessarily correlated only to long-distance flight. Birds with more aerial lifestyles, who’s survival depends on consistent flight, have also been shown to have high HWIs31. Therefore, it can be inferred that a high HWI is related to a high overall volume of flight, and evolution towards flight efficiency.
Some studies have connected the HWI more directly to flight efficiency. One study of swain’s thrush(Catharus ustulatus) found that individual birds with especially “pointy” wings exhibited lower heart rates in migratory flight33. A lower heart rate means a slower metabolism, which consumes less energy, making high AR wings more efficient. Other studies have found that birds with high HWI are able to fly further than those with low HWI, further suggesting that the HWI is linked to flight efficiency31. The connection between a high AR and decreased induced drag due to weaker wingtip vortices likely applies to high HWI as well. However, the aerodynamic complexities of bird flight hinder researchers from making conclusive observations.
The HWI has also been shown to increase with elevation, particularly at 4 km above sea level34. For flapping birds in particular, the increase of the HWI with elevation is particularly significant. In soaring birds, on the other hand, this effect is much less clear, possibly due to the fundamental aerodynamic differences between flapping and soaring flight. In addition, soaring birds are generally restricted to lower altitudes, which could further affect the change in their HWI34. This trend aligns with the higher ARs to decrease drag at higher altitudes, where the less air pressure leads to a decrease in lift.
Studies have also connected higher HWI to a bird’s ability to respond to wind, and the associated aerodynamic forces. With higher HWI wings, the migration-inhibiting effects of headwind seemed to decrease, indicating a higher HWI correlates to an increased ability to cope with wind and the drag associated with random airflow35. However, these studies remain fairly limited, and explanations for the correlation between HWI and wind effects have not been substantiated.
Flight and Reynolds numbers
Flight occurs across a wide range of scales and environments, which adds complexity when comparing organisms of different sizes. For example, insects and airplanes are vastly different in size and therefore operate in very different aerodynamic conditions. Reynolds number, a ratio of inertial to viscous forces in a fluid, is one of the most important parameters when comparing flight across different scales. As seen in the equation, the larger the characteristic dimension (or scale) of a flying body, the larger the Reynolds number. As Reynolds number increases, the flow shifts from laminar to turbulent. For flying organisms and vehicles, the wingspan or area is typically used as the characteristic length. From this, we can also see that Reynolds number generally increases as flow speed increases, meaning a body flying at a higher velocity will experience more turbulent flow.
(10) 
In environments with low Reynolds number, such as at low speeds or small scales, airflow tends to be more laminar. For high aspect ratio (AR) wings, this often results in a reduced ability to generate lift, and thus lower flight efficiency. In laminar flow, the fluid moves in smooth, parallel layers, making flow separation more likely. As a result, at low speeds or small scales, high AR wings are unable to maintain lift, leading to a loss in flight efficiency. Low AR wings, however, can be more efficient in low Reynolds number environments. With laminar flow, the air is typically more viscous and less inertial, which dampens vortex instabilities. For low AR wings, this leads to more stable leading-edge vortices (LEVs), generating additional lift and improving flight efficiency. High AR wings do not benefit as much from the stability of LEVs since they lack strong spanwise flow, causing their LEVs to detach more quickly and contribute less to lift. At low Reynolds numbers, where the flow is predominantly viscous and sticky, generating lift through gliding becomes nearly impossible. In these conditions, flapping is essential to maintain lift, which complicates the use of high AR wings, as they are typically more structurally fragile.
In environments with high Reynolds number, such as at higher speeds or larger scales, airflow is usually more turbulent. For high AR wings, this turbulent flow helps keep the flow attached, even when the wing is particularly thin or long, increasing lift while keeping drag low and improving overall flight efficiency. However, the increased turbulence also makes the wings more susceptible to bending, so the aspect ratio can only be increased to a certain extent before structural issues arise. At high Reynolds number, LEVs are more prone to breaking down, meaning low AR wings struggle to generate lift and lose flight efficiency. For high Reynolds number environments, where the flow is more inertial and attached, gliding becomes more viable, while the efficiency of flapping decreases.
Between low and high Reynolds numbers, the efficiency of high and low AR wings depends on factors such as shape, flapping speed, and other environmental conditions.
The variation in Reynolds number across scales plays a critical role in designing bio-inspired wings. While high AR wings are optimal for endurance and overall efficiency, they lose lift generation capabilities at small scales and low speeds. As such, they are better suited for high Reynolds number environments, such as those seen in large birds or even commercial airplanes. In contrast, low AR wings retain, and sometimes even gain, efficiency at low Reynolds numbers, making them more suitable for small-scale drones.
Morphing Wings
Nearly all flying animals, ranging from insects to birds to bats and beyond, morph their wings in flight. Studies on other organisms, such as gliding lizards, include important analyses as well, but are not included here36. Only organisms in long range flight, such as migratory birds, keep their wings rigid for extended periods of time. Even then, when forced to land or perch, their wings twist and morph beyond their original shape. Wing morphing can be largely split into three categories: variable planform, camber and twist11. This paper will mainly focus on changes in wing planform, which is influenced by the AR. During bird and insect flight, most changes in wing planform occur through flapping, and passive deformations due to wing flexibility. These morphings, however, are highly temporary, and do not represent true changes in the organism’s AR. Therefore, this paper will focus mainly on wing morphing in gliding, where changes are somewhat more permanent, and more directly reflect changes in the environment.
In Birds
Bird wings, which consist of feathers and skin layered over flexible bones and joints, are highly variable, constantly changing in flight. Typical bird wings consist of 10 different skeletal elements, much like a human hand, giving birds a large amount of control over the shape of their wings. Wing feathers are layered across the wing, and come in a range of sizes according to their function. As a result of how their feathers are layered, bird wing planforms can radically change, stretching and retracting according to environmental changes11.
While flapping, bird wings continually twist and bend, altering their planform. Such changes, however, do not necessarily present a true modification of the AR. The AR is a permanent measure of wing geometry; the ‘effective’ AR created by flapping flight is not held for long. Therefore, changes of the effective AR during flapping flight will not alter the long term aerodynamic performance of the wing significantly. However, some types of effective AR are more permanent, and do affect aerodynamic performance. In gliding flight, for example, birds morph their wings according to environmental conditions, and maintain those geometries for extended periods of time.
Birds use gliding flight to conserve energy, catching updrafts and utilizing their forward motion to generate lift, rather than constantly flapping. To optimize their efficiency, birds adjust their ARs in flight. Soaring gulls, for example, adjust their wing area, retracting their wings as wind increases, and unfolding them once again when they lose too much altitude5. While soaring, and catching updrafts, gulls need lower AR wings and higher wing areas to prevent stalling at low speeds and to brave the turbulent airflow. Gliding falcons perform the same maneuver. Tucker and Parrott observed a falco jugger, or lugger falcon, in a wind tunnel at various airspeeds, and calculated the bird’s wing span and area (Fig. 5). This study found that the falcon consistently approached the maximum L/D ratio at given airspeeds, by adjusting its wing span and wet area37.

(11) 
Where S is the wing area, and b is the wing span
(12) 
The aspect ratio, with the wing span and area from the previous equation.
As the airspeed was increased, the falcon retracted its wings into an ‘M’ shape, decreasing its wingspan. According to the least squares regression line calculated for the falcon’s wingspan and wing area, increasing the wingspan exponentially increased the AR as well. Therefore, the falcon also adjusted its AR to achieve the maximum possible L/D ratio, and flight efficiency. Harris hawks have also been shown to reduce their wing area by half, adjusting to an increase in airspeed11. Numeric simulations of a flapping wing in flight confirm these measurements, showing that dynamic changes of the wingspan, and therefore AR, greatly enhance the lift produced by a wing. Changing wingspans were shown to intensify LEVs on the upper surface of the wing, while also weakening the vortices on the lower surface that generate negative lift, therefore creating a much higher L/D ratio38.
While slowing down to perch, birds morph their wings as well. A steppe eagle, or aquila nipalensis, was repeatedly filmed while perching to document the morphing of its wing39. When highly flexed, bird wings adopted an ‘M’ shape, as seen in Figure 5. As the steppe eagle approaches to perch, it flexes its wings, altering their area and AR. This maneuver rapidly increases drag, and brings the bird’s center of pressure forward, leading to a pitch-up motion that adds to the drag. At the end of the pitch-up motion, the wings are stretched out again to their usual AR. At such slow speeds, however, this quickly leads to stall, allowing the bird to complete the perching maneuver39 This perching maneuver clearly illustrates how birds adjust their wing geometries, and AR, to aid their agility.
In diving, and other fast maneuvers, birds also radically change the AR of their wings. Falcons, especially peregrine falcons, are renowned for their stoops, or rapid dives to catch prey. It has been predicted that peregrine falcons may reach speeds over 80 meters per seconds during their stoops, and have the ability to attain even higher speeds40. When stooping, falcons first retract their wings to achieve high speeds, before extending them again as they approach their prey. In the most compact formation, known as the teardrop shape, the falcon folds its wings in as far as possible to reduce drag, allowing it to speed up. Then, its wings are extended back into an ‘M’ shape, increasing drag and helping the falcon eventually pull out of its dive. Between these shapes, however, falcons constantly change their wing shape, and effective AR, to control the rate of their descent and zero in on their target. Their stoop is a constant balance between lift and drag, controlled by morphing their wings and the Ars41.
In Insects
Unlike bird wings, insect wings are made of a membrane supported by veins, rather than overlapping feathers. Their wings are therefore largely unable to stretch, or retract, restricting changes in their wing planform. However, changes in the effective AR do still occur through twisting, and wing overlap while flapping.
Insects are typically very small, light, and fly at relatively slow speeds. Therefore, their wings must be light, while still holding up to the aerodynamic forces of being flapped, meaning their wings are often highly flexible. This means that most of the morphing in insect wings planform is passive resulting in the natural deformations of their wings rather than an active response to environmental conditions11. These changes are also highly temporary, gone again before the changes in AR fully take effect. On their upstroke, for example, insects fold their wings into their bodies, briefly diminishing their wing area and AR to decrease negative forces. This change does improve the efficiency of the insect’s flight, but not due to a change in the AR, but rather because of less wing area producing drag. The flexibility of insect wings also leads to benefits like the ‘umbrella effect’ in locusts. The umbrella effect occurs when the trailing edge of the wing flexes forward during the downstroke, leading to compressive forces in the veins that induce positive camber. This response helps the wing resist further deformations from upward forces, enhancing its structural stability, and lift generation. The umbrella effect is a passive result of the wing’s structure, rather than an active response by the insect to its environment, meaning it does not represent a true morphing of the insect’s AR.
Insects with multiple wings on each side can change the overlapping of their wings, and exhibit more permanent wing morphing. Like birds who change the configuration of their feathers, creating more overlap between multiple wings will also morph the overall wing planform11. However, this kind of morphing has not been extensively studied, and its impacts on flight are therefore unknown.
Morphing Wing Designs
Morphing Wing Designs
For the last 100 years, researchers around the world have been working on flight vehicles with morphing wings. In the US, NASA and DARPA have inspired and funded many research projects, particularly around smart memory alloys (SMAs) and flexible wings. Internationally, China and the EU have also contributed research and designs to this field, especially as the practicality and military applications of morphing wings became more widespread42. To date, most morphing wings have focused on a single degree of freedom, or type of wing morphing. However, some designs have begun incorporating several types of morphing, and therefore further approaching biomechanical flight.
Designs focusing specifically on changing the AR, or wingspan, have been present since the 1930s, though few have been flown. Many of these aircrafts use telescopic designs, in which portions of the wing slide into each other, allowing it to extend and retract. The first plane to fly with telescopic wings was the MAK – 10, designed by Ivan Makhonine, a Russian expatriate. Through pneumatic powering, MAK – 10 was able to increase its span by 62%, and its area by 57%, leading to a radical change in the AR from 8.05 to 13.443. Similar designs based off the MAK-10 in more recent years have achieved changes in the AR of up to 230%. Another design, the Akaflieg Stuttgart FS-29, was built in Germany from 1972. This plane achieved an ARs of 20.67 and 28.54, very high values that reflect its size and long-distance applications. Other designs with changes in the AR have emerged in more recent years, some of which are outlined below.
Lockheed Martin
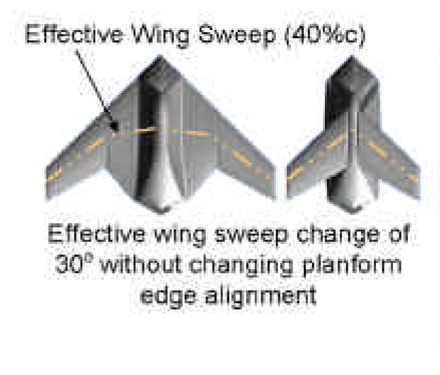
In the US, funding through DARPA led to several designs for flying vehicles with morphing wings. One such design created by Lockheed Martin Aerodynamics between 2003-2006 featured a wing with a telescopic mechanism (Fig. 6), essentially creating two modes of flight44. This drone was designed as a ‘Hunter-Killer,’ an example of the possible versatility of MAVs. In the hunting mode, the drone was meant to search, and prioritize flight efficiency to survey large areas. The killer mode, on the other hand, sought to increase speed and maneuverability to ‘kill’. The Lockheed design put particular emphasis on the wing sweep, which changes by 30°, wing area, and wetted area to increase flight efficiency and versatility. The morphing aspect of this wing also greatly increases the surface area, and somewhat increases the wing span. Calculations predict that compared to conventional flight vehicles, the Lockheed drone has a 22% higher mission radius, meaning it could fly further on less fuel44. However, designs for the conventional vehicles used were constrained by fuel storage in the wings, as the root chord needed to remain the same, meaning conventional vehicles may perform more efficiently than predicted.
According to the wing measurements given, the Lockheed design does not undergo a large change in AR. In its retracted state, the full-scale model is designed with a span of 24.1 feet and an area of 215.5 square feet, while the extended mode has a span of 215.5 feet and an area of 600 square feet. Therefore, the AR in the retracted state is roughly 2.7, and extends to an AR of 2.8444. Compared to the wide range of AR in birds, from 5.5 to 19, these values are very small, and the change between them is almost negligible. To further increase the efficiency of its flight, the Lockheed design would likely need a larger change in AR, perhaps through a second section of sliding wing area.
MX-1
Another design funded by DARPA was the MX-1, built by NextGen Aeronautics and flown in 2007 (Fig. 7). This design focused on a single degree of freedom, morphing the wing span and area. The final vehicle was much larger than other designs, weighing around 100 pounds with a wingspan of 9.3 feet. This design included flexible wing skins, capable of stretching as the wing morphs into a different shape. During flight, the wings successfully underwent a 40% change in wing area, and a 30% change in the wing span45. Using these percentages, we can estimate a 21% increase in the AR by morphing the wing, possibly helping to increase flight efficiency in the extended state. The two modes in this design are more similar to high and low AR wings in birds, including an overall increase in wing pointedness and a decrease in wing breadth.

LisEagle
The LisEagle is one of the most advanced biomechanically inspired drones to date, with feathered wings and tail that morph much like those on birds (Fig. 8). This drone did not focus on morphing the AR specifically – instead, it sought to imitate bird wings, and achieve several degrees of freedom. The LisEagle was initially built in 2020, and was an ongoing project through 202247,48. Through several interdependent actuators, this drone can morph its wing and tail shape, as well as twist its wings in response to environmental conditions. It can also perform perching maneuvers, such as a rapid pitch-up to increase drag49. By adapting to environmental changes, the LisEagle has exhibited stable flight despite disturbances, including physical nudges and turbulent airflow. Measurements of the LisEagle in flight suggest that by morphing its wings, it can achieve significantly increased energy efficiency, especially in long range flight. By retracting and extending its feathers, the LisEagle does exhibit some changes in its AR ratio as well, similar to the wing morphing present in gliding birds. However, it remains constrained to gliding and turning maneuvers, unable to fold in its wings as falcons do while stooping.

Morphing AR for Mechanical Flight
As established in the previous sections, both low and high AR wings have benefits. Both biological organisms and mechanical designers have realized morphing between the two as a method to increase flight efficiency.
Wings with an aspect ratio above 8 are ideal for long distance flight at high speeds, and at high altitudes, since they can generate lift with minimal drag. These wings are best for non-flapping applications, since their slender structure puts them at risk of bending under high pressure. Migratory birds, which fly long distances at high speeds and altitudes, often have wings with ARs between 4 and 85. Commercial airplanes have even higher ARs, with values around 8-10, and sailplane ARs can even reach into the 40s or 50s1. High AR wings would be ideal for the section of any mission that requires steady long-distance flight with few maneuvers, possibly at high altitudes. In the context of the aforementioned “hunter-killer” aircrafts, high AR wings would be very applicable to the ‘hunter’ portion of flight, where the aircraft surveils large areas for its target. More generally, high AR wings are applicable to any mission that requires surveillance across a large area, transport over long distances, or even flight at high altitudes. Using high AR wings would help manage fuel usage for missions like these, and saving power for other sections of the mission.
Low AR wings, within the range of 2.5-6, are well suited for short distance, agile flight, at slow speeds and low altitudes, because they stall at much slower speeds and can easily recover from losing lift. these wings are very applicable to flapping, especially because of the small-scale environments they usually exist in. Small, agile birds, capable for performing complex dives and turns, often have wings with ARs around 3-45. Larger aircrafts that carry out fast maneuvers, like fighter jets, often also have low AR wings even though they exist in high Re environments. Low AR wings would be ideal for any mission that requires fast and exact turns, at low or high altitudes, usually at somewhat smaller scales. They would be perfect for the ‘killer’ portion of ‘hunter-killer’ aircrafts, helping the vehicle execute fast turns and fly at low speeds to meet its target. Other applications include small-scale drones and indoor flying vehicles, both of which would benefit from a high degree of agility.
Pairing low and high AR wings together allows flight vehicles to take on multiple different modes, and save fuel overall. Morphing between the two, an aircraft could surveil large areas using relatively little fuel, and then execute fast maneuvers or fly at slower speeds for closer inspection when necessary.
Discussion
The AR is an important parameter of wing shape, widely used in traditional equations for flight design and in research into biomechanical flight. In airplane design, a high AR correlates to low induced drag, and increased flight efficiency. In contrast, low ARs typically represent high maneuverability, and better performance at slow speeds due to supporting higher AOA. These trends are largely mirrored by research on bird flight, where high AR are associated with long-distance migration and high-altitude flight3,6,16,20. Particularly agile birds also utilize low ARs to achieve fast speeds and turns6,21. In bird flight, the AR ratio is highly correlated to the HWI, a more accessible measurement29. The HWI in birds largely corroborates trends between the AR and flight efficiency30,31 ,32 ,33 ,34,35. In insects, these trends are much less clear, although some evidence supports a link between high AR and flight efficiency22,26 ,24 ,51. Both birds and insects morph their wings during flight, although most of this morphing occurs passively10. Designs for aircrafts with morphing wings have emerged globally in the last century, though there have been few actual flights of vehicles with significantly changing ARs44,52 ,47,48. Those that have been flown are restricted to a single kind of morphing, changing only the wing planform rather than utilizing changes in camber and twist seen in other designs.
This paper highlights the importance of the AR in wing design, bridging the gap between engineering models and biological examples. By considering the AR across multiple disciplines, we can look for broader trends confirmed by more than one kind of flight. Traditional equations, while useful in predicting general trends in flight efficiency and maneuverability, only consider rigid, non-flapping wings in idealized flight. Studies of plane flight have some of the same limitations, by including only stationary, gliding wings. By incorporating insights from biological systems, we can expand our knowledge of how ARs affect flight in real world conditions, and under different types of flight. Bird flight in particular includes a wide range of styles, from flapping to gliding and combinations of the two, confirming the effects of the AR beyond stationary wings5. Knowing that trends in the AR remain largely true in both stationary and flapping flight highlights the importance of wings with morphing ARs in future designs.
In addition, understanding the limitations of morphing in bird and insect flight helps to inform future morphing wing designs. Though biomechanical wings often exhibit some level of morphing, through changes in planform, camber, and twist, there are very few examples of significant changes in the AR. Insects in particular exhibit very little morphing of the AR, mainly adapting their wings through camber and twist11. Birds, who have overlapping feathers, can actively morph their wings while gliding and diving, although significant changes in AR are still restricted38,39,40,41. Very few birds are able to change their ARs radically enough to exhibit both high flight efficiency, and high maneuverability, through changes in their wings. Therefore, future wing designs will need to consider morphing technologies beyond those seen in birds and insects, to fully obtain the benefits of both high and low ARs.
Lastly, the development of new mechanisms for altering the AR is necessary, to create wings capable of changing both the AR and other parameters of flight. While the AR is an important parameter of flight, connected to the wing span and wing area, achieving maximum flight efficiency is only possible through other kinds of wing morphing as well. Current AR morphing designs, such as the MX-1 or Lockheed, are restricted to changing the wing planform, and are unable to alter the twist or camber of their wings as other designs can44,52. Therefore, future designs will need to incorporate changes in the AR, without neglecting other kinds of morphing.
Limitations
While the AR is an important parameter of wing shape, it affects only a section of the lift and drag produced in flight. For example, while the AR affects the LEV and induced drag, it does not address other key components of flight such as wing flexibility, camber, or the AOA. The AR is also blind to 3D changes in wing shape, such as the C-shape adopted by stooping birds, and therefore cannot fully account for the all the dynamic adjustments in wing morphology that contribute to flight performance. In addition, the AR doesn’t consider the role of other flight surfaces in generating aerodynamic forces. In birds the tail plays a large role in generating forces, granting stability and steering capabilities10. Morphing the tail allows for active adaptations to environmental changes in flight, which the AR does not measure. Though the AR does affect flight, other parameters can lead to high levels of flight efficiency and maneuverability that aren’t reflected in this measurement.
Conclusion
This study examines the influence of different aspect ratios (ARs) on flight efficiency across biological and mechanical systems. It concludes that high AR wings are best suited for high-speed, long-distance flight, while low AR wings excel in maneuverability. These patterns are consistently observed in both mechanical flight and bird flight, though they are less well-defined in insect flight research. The connection between AR and performance was further explored through the lens of biological and mechanical morphing wings, which can switch between flight modes by altering their ARs.
Despite this, biological systems like birds and insects exhibit limited ability to significantly change their ARs mid-flight. Meanwhile, current mechanical morphing wing designs remain largely restricted to changes in planform alone. To fully capitalize on the benefits of both high and low ARs, future wing designs must incorporate more advanced morphing capabilities that extend beyond planform adjustments. This includes the integration of changes in camber, twist, and other aerodynamic features. By drawing inspiration from both engineered flight systems and natural flyers, future morphing wing technologies can push the boundaries of flight versatility and efficiency across diverse conditions.
References
- Anderson JD. Fundamentals of Aerodynamics. McGraw-Hill Education; 2017. 1130 p. [↩] [↩] [↩] [↩] [↩] [↩]
- Gudmundsson S. Chapter 9 – The Anatomy of the Wing. In: Gudmundsson S, editor. General Aviation Aircraft Design [Internet]. Boston: Butterworth-Heinemann; 2014 [cited 2024 Dec 11]. p. 299–399. Available from: https://www.sciencedirect.com/science/article/pii/B978012397308500009X [↩]
- Claramunt S. Flight efficiency explains differences in natal dispersal distances in birds. Ecology. 2021;102(9):e03442. [↩] [↩] [↩] [↩]
- Taylor GK, Thomas ALR. Evolutionary Biomechanics: Selection, Phylogeny, and Constraint. Oxford University Press; 2014. 172 p. [↩] [↩]
- Tennekes H. The Simple Science of Flight, revised and expanded edition: From Insects to Jumbo Jets. MIT Press; 2009. 217 p. [↩] [↩] [↩] [↩] [↩] [↩] [↩] [↩] [↩] [↩]
- Lockwood R, Swaddle JP, Rayner JMV. Avian Wingtip Shape Reconsidered: Wingtip Shape Indices and Morphological Adaptations to Migration. J Avian Biol. 1998;29(3):273–92. [↩] [↩] [↩] [↩] [↩] [↩] [↩] [↩]
- Potty V, Angelo S, Rao PS, G S. Recent Developments of an Aircraft Fuselage along Theoretical, Experimental and Numerical Approach – A Review. Univers J Mech Eng. 2019 Dec;7(6A):21–8. [↩]
-
Tennekes H. The Simple Science of Flight, revised and expanded edition: From Insects to Jumbo Jets. MIT Press; 2009. 217 p. [↩]
- Alerstam T. Bird f light and optimal migration. Trends Ecol Evol. 1991 Jul 1;6(7):210–5. [↩]
- Shyy W, Berg M, Ljungqvist D. Flapping and flexible wings for biological and micro air vehicles. Prog Aerosp Sci. 1999 Jul 1;35(5):455–505. [↩] [↩] [↩]
- Taylor GK, Carruthers AC, Hubel TY, Walker SM. Wing Morphing in Insects, Birds and Bats: Mechanism and Function. In: Morphing Aerospace Vehicles and Structures [Internet]. John Wiley & Sons, Ltd; 2012 [cited 2024 Dec 13]. p. 11–40. Available from: https://onlinelibrary.wiley.com/doi/abs/10.1002/9781119964032.ch2 [↩] [↩] [↩] [↩] [↩] [↩] [↩]
- Heathcote S, Wang Z, Gursul I. Effect of spanwise flexibility on flapping wing propulsion. J Fluids Struct. 2008 Feb 1;24(2):183–99. [↩]
- Landell-Mills N. Insect flight explained according to Newtonian physics. [Internet]. 2024 [cited 2024 Dec 11]. Available from: https://www.researchgate.net/publication/336312526_Insect_flight_explained_according_to_Newtonian_physics [↩] [↩]
- Munson BR, Rothmayer AP, Okiishi TH. Fundamentals of Fluid Mechanics, 7th Edition. Wiley; 2012. 796 p. [↩]
- Gudmundsson S. Chapter 9 – The Anatomy of the Wing. In: Gudmundsson S, editor. General Aviation Aircraft Design [Internet]. Boston: Butterworth-Heinemann; 2014 [cited 2024 Dec 11]. p. 299–399. Available from: https://www.sciencedirect.com/science/article/pii/B978012397308500009X [↩]
- Minias P, Meissner W, Włodarczyk R, Ożarowska A, Piasecka A, Kaczmarek K, et al. Wing shape and migration in shorebirds: a comparative study. Ibis. 2015;157(3):528–35. [↩] [↩]
- ie C, Gao N, Meng Y, Wu Y, Yang C. A review of bird-like flapping wing with high aspect ratio. Chin J Aeronaut. 2023 Jan 1;36(1):22–44. [↩]
- Kruyt JW, Quicazán-Rubio EM, van Heijst GF, Altshuler DL, Lentink D. Hummingbird wing efficacy depends on aspect ratio and compares with helicopter rotors. J R Soc Interface. 2014 Oct 6;11(99):20140585. [↩]
- Rayner JMV. Form and Function in Avian Flight. In: Johnston RF, editor. Current Ornithology [Internet]. Boston, MA: Springer US; 1988 [cited 2024 Dec 11]. p. 1–66. Available from: https://doi.org/10.1007/978-1-4615-6787-5_1 [↩]
- Xie C, Gao N, Meng Y, Wu Y, Yang C. A review of bird-like flapping wing with high aspect ratio. Chin J Aeronaut. 2023 Jan 1;36(1):22–44. [↩] [↩]
- van den Hout PJ, Mathot KJ, Maas LRM, Piersma T. Predator escape tactics in birds: linking ecology and aerodynamics. Behav Ecol. 2010 Jan 1;21(1):16–25. [↩] [↩]
- Bhat SS, Zhao J(赵季生), Sheridan J, Hourigan K, Thompson MC. Aspect ratio studies on insect wings. Phys Fluids. 2019 Dec 19;31(12):121301. [↩] [↩] [↩] [↩] [↩]
- N, Knowles K, Bomphrey RJ. The effect of aspect ratio on the leading-edge vortex over an insect-like flapping wing. Bioinspir Biomim. 2015 Oct 9;10(5):056020. [↩]
- Broadley P, Nabawy MR, Quinn MK, Crowther WJ. Wing planform effects on the aerodynamic performance of insect-like revolving wings. In: AIAA AVIATION 2020 FORUM [Internet]. American Institute of Aeronautics and Astronautics; 2020 [cited 2024 Dec 11]. (AIAA AVIATION Forum). Available from: https://arc.aiaa.org/doi/10.2514/6.2020-2667 [↩] [↩] [↩]
- Harbig RR, Sheridan J, Thompson MC. Reynolds number and aspect ratio effects on the leading-edge vortex for rotating insect wing planforms. J Fluid Mech. 2013 Feb;717:166–92. [↩] [↩] [↩]
- Phillips N, Knowles K, Bomphrey RJ. The effect of aspect ratio on the leading-edge vortex over an insect-like flapping wing. Bioinspir Biomim. 2015 Oct 9;10(5):056020. [↩] [↩] [↩] [↩]
- Chu JJ, Claramunt S. Determinants of natal dispersal distances in North American birds. Ecol Evol. 2023 Feb 8;13(2):e9789. [↩]
- Claramunt S, Wright NA. Using Museum Specimens to Study Flight and Dispersal1. In: The Extended Specimen. CRC Press; 2017. [↩]
- Claramunt S, Derryberry EP, Remsen JV, Brumfield RT. High dispersal ability inhibits speciation in a continental radiation of passerine birds. Proc Biol Sci. 2012;279(1733):1567–74. [↩] [↩] [↩] [↩]
- Sheard C, Neate-Clegg MHC, Alioravainen N, Jones SEI, Vincent C, MacGregor HEA, et al. Ecological drivers of global gradients in avian dispersal inferred from wing morphology. Nat Commun. 2020 May 18;11(1):2463. [↩] [↩] [↩]
- Weeks BC, O’Brien BK, Chu JJ, Claramunt S, Sheard C, Tobias JA. Morphological adaptations linked to flight efficiency and aerial lifestyle determine natal dispersal distance in birds. Funct Ecol. 2022;36(7):1681–9. [↩] [↩] [↩] [↩]
- Dawideit BA, Phillimore AB, Laube I, Leisler B, Böhning-Gaese K. Ecomorphological predictors of natal dispersal distances in birds. J Anim Ecol. 2009;78(2):388–95. [↩] [↩]
- Bowlin MS, Wikelski M. Pointed Wings, Low Wingloading and Calm Air Reduce Migratory Flight Costs in Songbirds. PLOS ONE. 2008 May 14;3(5):e2154. [↩] [↩]
- Yang J, Yang C, Lin H wei, Lees AC, Tobias JA. Elevational constraints on flight efficiency shape global gradients in avian wing morphology [Internet]. bioRxiv; 2024 [cited 2024 Dec 11]. p. 2024.07.12.603304. Available from: https://www.biorxiv.org/content/10.1101/2024.07.12.603304v1 [↩] [↩] [↩]
- Laske V, Immelman K. Eine Beziehung zwischen Handflügelindex and Windempfindlichkeit bei Zugvögeln. Volgelwarte. 31:177–8. [↩] [↩]
- Khandelwal PC, Hedrick TL. Combined effects of body posture and three-dimensional wing shape enable efficient gliding in flying lizards. Sci Rep. 2022 Feb 2;12(1):1793. [↩]
- Tucker VA, Parrott GC. Aerodynamics of Gliding Flight in A Falcon and Other Birds. J Exp Biol. 1970 Apr 1;52(2):345–67. [↩] [↩]
- Wang S, Zhang X, He G, Liu T. Lift enhancement by dynamically changing wingspan in forward flapping flight. Phys Fluids. 2014 Jun 24;26(6):061903. [↩] [↩]
- Carruthers AC, Thomas ALR, Taylor GK. Automatic aeroelastic devices in the wings of a steppe eagle Aquila nipalensis. J Exp Biol. 2007 Dec;210(Pt 23):4136–49. [↩] [↩] [↩]
- Hedenström A. The stoop of large falcons. Trends Ecol Evol. 1998 Oct 1;13(10):383–5. [↩] [↩]
- Gowree ER, Jagadeesh C, Talboys E, Lagemann C, Brücker C. Vortices enable the complex aerobatics of peregrine falcons. Commun Biol. 2018 Apr 5;1(1):1–7. [↩] [↩]
- Chu L, Li Q, Gu F, Du X, He Y, Deng Y. Design, modeling, and control of morphing aircraft: A review. Chin J Aeronaut. 2022 May 1;35(5):220–46. [↩]
- Ajaj RM, Saavedra Flores EI, Friswell MI, De la O FAD. Span Morphing Using the Compliant Spar. J Aerosp Eng. 2015 Jul 1;28(4):04014108. [↩]
- Bye D, McClure P. Design of a morphing vehicle. 48th AIAA/ASME/ASCE/AHS/ASC Structures, Structural Dynamics, and Materials Conference. 2007;:1–8. [↩] [↩] [↩] [↩] [↩] [↩]
- Flanagan J, Strutzenberg R, Myers R, Rodrian J. Development and flight testing of a morphing aircraft, the NextGen MFX-1. 48th AIAA/ASME/ASCE/AHS/ASC Structures, Structural Dynamics, and Materials Conference. 2007;:1–10. [↩]
- Flanagan J, Strutzenberg R, Myers R, Rodrian J. Development and flight testing of a morphing aircraft, the NextGen MFX-1. 48th AIAA/ASME/ASCE/AHS/ASC Structures, Structural Dynamics, and Materials Conference. 2007;:1–10. [↩]
- Ajanic E, Feroskhan M, Mintchev S, Noca F, Floreano D. Bioinspired wing and tail morphing extends drone flight capabilities. Sci Robot. 2020 Oct 28;5(47):eabc2897. [↩] [↩]
- Ajanic E, Feroskhan M, Wüest V, Floreano D. Sharp turning maneuvers with avian-inspired wing and tail morphing. Commun Eng. 2022 Nov 24;1(1):1–9. [↩] [↩]
- Wüest V, Jeger S, Feroskhan M, Ajanic E, Bergonti F, Floreano D. Agile perching maneuvers in birds and morphing-wing drones. Nat Commun. 2024 Sep 27;15(1):8330. [↩]
- Jeger SL, Wüest V, Toumieh C, Floreano D. Adaptive morphing of wing and tail for stable, resilient, and energy-efficient flight of avian-inspired drones. Npj Robot. 2024 Nov 20;2(1):1–10. [↩]
- Harbig RR, Sheridan J, Thompson MC. Reynolds number and aspect ratio effects on the leading-edge vortex for rotating insect wing planforms. J Fluid Mech. 2013 Feb;717:166–92 [↩]
- Flanagan J, Strutzenberg R, Myers R, Rodrian J. Development and flight testing of a morphing aircraft, the NextGen MFX-1. 48th AIAA/ASME/ASCE/AHS/ASC Structures, Structural Dynamics, and Materials Conference. 2007;:1–10. [↩] [↩]


