
Abstract
This study examined associations between probiotic supplementation and changes in the human gut microbiota, along with microbial responses in Hermetia illucens larvae, to explore a low-cost and ethical alternative to vertebrate models. Ten participants consumed personalized probiotic supplements for four weeks, and stool samples were analyzed before and after the intervention using 16S rRNA sequencing. Based on the laboratory’s proprietary gut health grading scale, 7 of 10 participants showed increased scores, 1 showed no change, and 2 showed lower scores. Separately, 7 participants showed an increase in at least one probiotic-associated genus, including Lactobacillus and Bifidobacterium. As a small-scale pilot study, these findings suggest that probiotic intake is associated with changes in gut microbiota profiles. In a parallel experiment, Hermetia illucens larvae were exposed to probiotics through spraying at two concentrations. Increased total culturable bacterial counts under aerobic LB and TSA culture conditions were observed in the 1:10 concentration group compared to the control group. Because the larval analysis was based on CFU measurements under aerobic conditions, the results reflect total culturable bacterial load rather than microbial diversity or strain-specific colonization. Because the two systems were evaluated using fundamentally different methods, direct comparison between species is not appropriate. These findings suggest that Hermetia illucens larvae may serve as a practical model for exploratory microbiota research. However, due to the small sample size and uncontrolled design, causal effects cannot be established.
Keywords: Probiotics, gut microbiota, 16S rRNA sequencing, Hermetia illucens, culturable bacteria, pilot study, health behavior
Introduction
Human health depends heavily on the gut microbiota, a vital component of digestion and immune function. It also plays a key role in immune regulation and overall systemic health1,2. Dysbiosis has been associated with gastrointestinal disorders such as inflammatory bowel disease and metabolic conditions2,3,4.
Probiotics are live microorganisms that are thought to help maintain or restore balance in the gut microbiota5. Previous studies suggest that key probiotic species, including Lactobacillus and Bifidobacterium, may contribute to digestion and overall gut health5.
The human gut contains many different types of microbes, including bacteria, viruses, fungi, protozoa, and archaea. These microbes are known to contribute to host defense against harmful bacteria and to the regulation of immune responses1. They also send signals to the brain through a pathway called the gut–brain axis6. This connection has been suggested to influence not only digestion but also the immune system and brain function6. When the natural microbial balance is disrupted, a condition known as dysbiosis occurs. Dysbiosis has been linked to health problems such as inflammatory bowel disease and metabolic conditions2,3,4. One of the most common conditions related to microbiota imbalance is irritable bowel syndrome (IBS), which is characterized by symptoms such as abdominal discomfort, bloating, and irregular bowel movements7.
The “gut–oral axis” is an emerging concept suggesting that microbial interactions may extend across different body sites, including potential links between the oral cavity and the gastrointestinal tract8,9.
Previous research has indicated that microbial communities in different body regions may be interconnected8,9. However, the present study focuses specifically on gut microbiota responses to probiotic supplementation and does not directly assess oral microbiota or oral health outcomes.
Traditionally, researchers have used vertebrate animals (like mice) to study the microbiota. However, these models can involve ethical concerns and have motivated the development of alternative experimental approaches10. In contrast, Hermetia illucens larvae (black soldier fly larvae) offer a simpler and more ethical alternative. They are easy to maintain and have a gut microbiota influenced by environmental factors such as diet, making them useful for studying microbial responses in controlled conditions11,12. These models help researchers study host–microbe interactions in simplified biological systems13.
Despite these advantages, fewer studies have examined how external factors such as diet or environmental conditions influence insect microbiota12,14. This suggests that these models may be better suited for exploratory microbiota research rather than direct translation to human health. The present study aimed to examine microbial responses to probiotic use in both humans and Hermetia illucens larvae under different experimental conditions. This study proposes a small-scale, ethical model to examine how probiotics may influence different organisms. Because Hermetia illucens larvae are inexpensive to rear at scale, they may serve as a useful screening model for early-stage microbiota experiments. Given the exploratory nature of this work, the findings do not establish a causal relationship.
In this context, “gut microbiota” refers to the bacterial composition identified through 16S rRNA sequencing, whereas “microbiome” refers to the broader functional and genomic context of these microbial communities.
Methods
Research Design
This study used a parallel experimental design to evaluate the effects of probiotic supplementation on gut microbiota using both human participants and an insect model (Hermetia illucens larvae). A before-and-after approach was used for human subjects, while a controlled laboratory experiment with multiple treatment groups was conducted for the insect model. This two-part design allowed microbial responses to probiotic exposure to be examined in both humans and Hermetia illucens larvae. Because the two experiments differed in exposure route, duration, and measurement method, the findings were interpreted separately.
Participants
Human Participants
A total of ten participants (5 males, 5 females; aged 13–78 years) were recruited. All participants were free from chronic illnesses, had not used antibiotics within the past year, and had no known gut-altering medical conditions. All volunteers participated willingly and were provided with detailed information regarding the study’s purpose and procedures.
Hermetia illucens Larvae
The insect experiment used 100 Hermetia illucens larvae, which were equally assigned into four groups (25 larvae per group). Each group’s larvae were pooled to form a single sample for CFU analysis (thus no independent replicates were obtained). The larvae were selected at a similar developmental stage to minimize biological variability.
Data Collection
Human Gut Microbiota Sampling
Participants collected fecal samples using the Greenbiome Gut kit (GC Genome) under sterile conditions. They swabbed the sample, placed it in the collection tube, stored it in a DNA buffer at room temperature, and sent it back within 24 hours using a courier service. Samples were analyzed using 16S rRNA sequencing to evaluate microbial diversity and bacterial abundance. The laboratory then calculated microbial health indices based on these results.
The gut microbiota score (grades F–S) was provided by the commercial laboratory based on its analysis of 16S rRNA sequencing data. The grading system reflects an overall evaluation of gut microbiota composition, including factors such as microbial diversity, balance, and selected health-related indicators. Scores are calculated based on multiple weighted components and converted into letter grades (F–S). Lower grades (e.g., F–C) indicate less balanced or less diverse microbial profiles, while higher grades (e.g., A–S) indicate relatively more balanced and diverse microbiota.
While the general basis of the scoring system is described, the detailed algorithm used to calculate the scores is proprietary and was not disclosed by the provider.
For visualization purposes, letter grades were converted into ordinal integers (F=1, C=2, B=3, A=4, S=5); no interval-scale meaning was assumed.
In addition to microbiota data, participants reported changes in symptoms such as bloating, digestive discomfort, fatigue, and bowel regularity. These observations were collected through self-report and were not assessed using standardized instruments. They therefore provide descriptive observations rather than quantitative clinical measures, and their interpretation is limited.
Larval Microbiota Sampling
For the insect model, a probiotic solution containing Lactobacillus and Bifidobacterium strains was applied directly to larvae through sterile spraying. The experiment consisted of four groups: Group A (baseline), Group B (control), Group C (1:10 probiotic), and Group D (1:1000 probiotic). Group A (baseline) was collected and processed at day 0, prior to any treatment. Group B received a sterile saline spray for five days. Group C received a 1:10 diluted probiotic spray applied three times daily, while Group D received a 1:1000 diluted probiotic spray under the same conditions.
Larvae were kept in plastic containers at a consistent density under general laboratory conditions. Temperature, humidity, and diet were not strictly controlled or systematically recorded. No dietary restrictions or fasting protocols were applied before sample processing.
Variables and Measurements
The main variable was probiotic exposure, either through capsule intake (humans) or spraying (larvae). Outcomes were assessed based on changes in microbiota composition in humans and levels of culturable bacteria in larvae, using 16S rRNA sequencing for humans and colony-forming unit (CFU) counts for larvae.
Human gut microbiota profiling targeted the V3–V4 hypervariable regions of the 16S rRNA gene using standard microbiota testing methods. Larval bacterial abundance was quantified using culture-based CFU counting.
Procedure
Human Experiment
Participants followed the sampling procedures described above using the Greenbiome Gut kit. The probiotic supplements were provided as commercially prepared capsules, either single-strain or multi-strain formulations, primarily including Lactobacillus and/or Bifidobacterium. The type of probiotic given to each participant was based on their initial microbiota results, although the exact criteria for this selection were not specified. The exact strain-level composition, CFU per dose, and manufacturer details were not disclosed by the provider. Participants were instructed to take the probiotic once daily throughout the study period. Compliance was monitored through daily self-report check-ins.
Diet and daily habits were not strictly controlled during the study. Participants were asked to keep their usual routines and avoid introducing new probiotic supplements or noticeably increasing fermented foods such as yogurt or kimchi. We did not use formal tools like food diaries or questionnaires. Instead, this aspect was checked informally through participant self-report. As a result, differences in diet or lifestyle may have influenced the findings.
Samples were then processed using 16S rRNA sequencing to assess microbial diversity and abundance, generating microbial health indices.
Larval Experiment
Following probiotic treatment, larvae were processed using a consistent protocol. All larvae in each group were anesthetized, surface-sterilized with 70% ethanol, and then homogenized in a buffer. The homogenized samples were diluted in six 10-fold steps (10⁻¹ to 10⁻⁶). From each dilution, 100 µL of the sample was plated onto agar plates for bacterial growth.
Data Analysis
We manually counted colony-forming units for each group. We analyzed the larval results descriptively by comparing CFU counts across experimental groups. Given the exploratory design and small sample size, we described the results without formal statistical tests.

Ethical Considerations
This study was conducted as an independent student research project under appropriate supervision and without formal Institutional Review Board (IRB) oversight. Informed consent was obtained from all participants, and parental or guardian consent was obtained for minors. All data were anonymized and managed in a way that did not allow identification of individual participants.
Results
Changes in Human Gut Microbiota
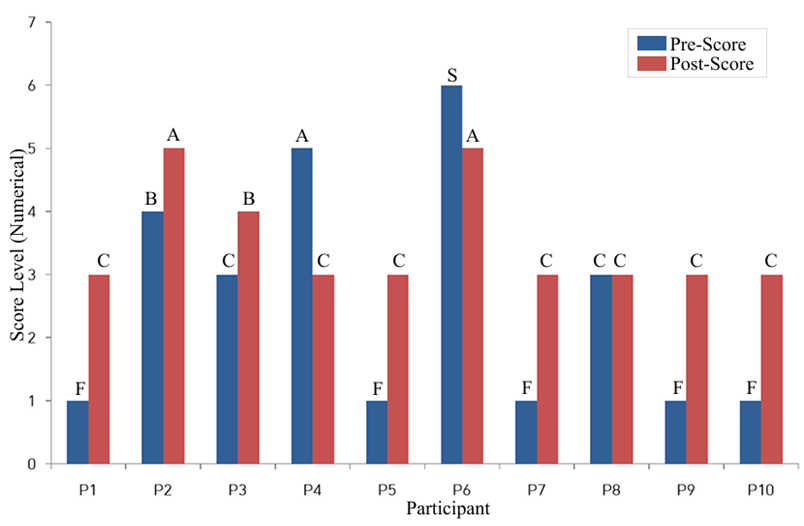
Based on the laboratory’s overall gut health grading scale, 7 of 10 participants showed an increase in post-intervention scores. One participant showed no change, and 2 participants showed lower scores. For example, Participant 1’s score changed from F to C, and Participant 2’s score changed from B to A, whereas Participant 4 changed from A to C and Participant 6 changed from S to A.
Separately, 7 of 10 participants exhibited an increase in the abundance of at least one probiotic-associated genus, including Lactobacillus or Bifidobacterium. However, this genus-level trend did not always match the overall grade outcome. Therefore, overall grade change was used as the primary outcome measure, while genus-level changes were reported separately.
Increases in Lactobacillus and Bifidobacterium were observed in several post-intervention summaries; these genera are commonly included in probiotic formulations and are widely associated with gut health5.
| Participant | Age (years) | Gender | Pre-Score | Post-Score | Probiotic Formula | Microbial Change |
| P1 | 78 | M | F | C | Lactobacillus Bifidobacterium | Lactobacillus (increase) Bifidobacterium (increase) |
| P2 | 74 | F | B | A | Bifidobacterium (only) | Bifidobacterium (increase) |
| P3 | 52 | M | C | B | Lactobacillus Bifidobacterium | Both (increase) |
| P4 | 48 | F | A | C | Lactobacillus (only) | Lactobacillus (decrease) |
| P5 | 47 | M | F | C | Lactobacillus Bifidobacterium | Both (increase) |
| P6 | 47 | F | S | A | Multi-strain blend | Overall diversity (decrease) |
| P7 | 19 | M | F | C | Lactobacillus (only) | Lactobacillus (increase) |
| P8 | 16 | M | C | C | Bifidobacterium (only) | No clear change |
| P9 | 16 | F | F | C | Bifidobacterium (only) | Bifidobacterium (increase) |
| P10 | 13 | M | F | C | Lactobacillus Bifidobacterium | Both (increase) |
The grading system used for these scores is described in Table 2.
| Grade | S | A | B | C | F |
| Score Range | 50–64 | 45–49 | 40–44 | 35–39 | ≤34 |
| Interpretation | Highest | High | Moderate | Low | Poor |
The grading system and score ranges are based on information provided in the laboratory report
Microbial Growth in Hermetia illucens Larvae
Groups B–D showed higher CFU counts than the baseline group A, with the highest values in Group C (1:10 dilution).
| Group | Treatment | LB (CFU/mL) | TSA (CFU/mL) |
| A | No treatment | 3.0 x 105 | 1.2 x 105 |
| B | Saline only | 5.5 x 106 | 3.0 x 106 |
| C | 10x probiotic | 7.2 x 106 | 4.3 x 106 |
| D | 1000x probiotic | 3.0 x 106 | 2.4 x 106 |
Total culturable bacterial counts under the aerobic culture conditions used in this study increased relative to the baseline group A, with the highest values observed in Group C. These measurements reflect only culturable bacteria under specific laboratory conditions and do not indicate microbial diversity or strain-specific colonization.
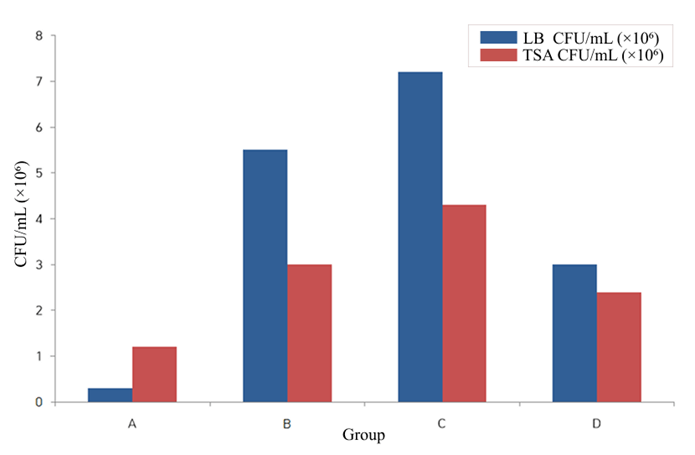
Figure 4 | Colony-forming unit (CFU/mL) counts of Hermetia illucens larvae across experimental groups A–D measured on LB and TSA media. Groups B–D showed higher CFU counts than the baseline group A, with Group C showing the highest values.
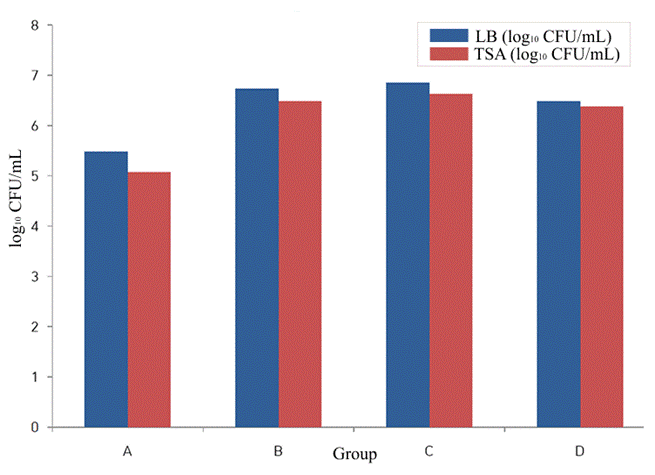
To better visualize relative differences across groups, the data were also log₁₀-transformed and are presented in Figure 5.
Summary of Experimental Outcomes
Both the human and insect experiments showed changes following probiotic exposure, although these changes were measured using different methods. In humans, these changes were reflected in microbiota scores and genus-level patterns. In larvae, changes in culturable bacterial counts were observed, with increases seen relative to the baseline group, although this pattern varied across treatment groups.
Discussion
Restatement of Key Findings
Probiotic supplementation was associated with changes in gut microbiota profiles. In Hermetia illucens larvae, increases in total culturable bacterial counts under the aerobic culture conditions used were observed in both probiotic-treated and control groups. Therefore, these findings should not be interpreted as changes in microbial diversity or gut microbiota composition. After four weeks of probiotic intake, seven out of ten human participants showed increased microbiota scores based on the laboratory’s grading scale, while one participant showed no change and two showed lower scores. Separately, seven participants showed increases in at least one of these genera, including Lactobacillus and Bifidobacterium, which have been associated with probiotic-related functions in the gut5. Some participants also reported reduced bloating and more regular digestion, but these observations were based on informal self-report and were not measured using standardized tools. Therefore, they should be considered as preliminary observations rather than formal clinical outcomes. Because the scoring system is proprietary, the biological meaning of specific grade changes could not be clearly determined.
In the insect model, increased culturable bacterial counts were observed with the highest values at the 1:10 probiotic concentration, while this pattern was not consistent across all treatment groups. Although the control group also showed an increase relative to baseline, the pattern suggests that factors other than probiotic concentration may have influenced bacterial growth. However, the human and larval experiments differed substantially in exposure route, duration, and measurement method. For this reason, the two sets of findings are best viewed independently, and any cross-species connection remains exploratory. This is consistent with previous studies showing that the gut microbiota of Hermetia illucens larvae varies depending on environmental conditions such as diet11,12.
Implications and Significance
Beyond digestive outcomes, several participants reported reduced fatigue and constipation. Although these observations are self-reported and were not assessed using standardized instruments, they may represent preliminary trends rather than confirmed physiological effects. We did not measure neuroendocrine or immune biomarkers; therefore, literature on the microbiota–gut–brain and gut–immune axes is provided here only as contextual background. Prior studies suggest that microbial activity can influence mood and other physiological systems6,15,16. These findings remain exploratory, and further studies using standardized measures and biological markers are needed to clarify these associations.
Insect models may be useful for early-stage exploratory research on probiotic-related bacterial responses.
Connection to Research Objectives
This work aimed to examine microbial responses to probiotic exposure in both humans and an insect model. Human participants showed changes in microbiota scores, while the larvae exhibited increases in culturable bacterial levels. Because the two experiments used different intervention methods and outcome measures, the findings do not support a direct cross-species comparison. However, the larval model may still be useful for exploratory investigation of probiotic-related microbial responses.
These findings are consistent with prior studies showing that probiotics have been widely studied for their roles in gut health and microbial balance17. However, individual-specific factors can influence how effectively probiotics colonize the gut, even among people consuming the same supplement. Notably, some participants showed limited response to probiotic supplementation. Such variability has also been observed in previous research, suggesting that probiotic effects can differ depending on individual factors18. In particular, a subset of participants did not show clear changes, suggesting that baseline microbiota composition may influence probiotic effectiveness. This supports the importance of personalized approaches to probiotic use.
Broader Implications for Health and Sustainability
Although oral microbiota or oral health outcomes were not directly assessed, recent research suggests a link between gut and oral health, with oral bacteria potentially contributing to gastrointestinal conditions such as inflammatory bowel disease8,9. For example, Fusobacterium nucleatum has been detected in colorectal tumors and has been associated with inflammation and tumor development19. These observations suggest that oral microbes may have broader systemic relevance, although this relationship remains indirect and was not directly tested.
In addition, Hermetia illucens has applications beyond microbiota research. It is used in processes such as the conversion of organic waste into biomass and the production of insect-based protein20. These characteristics suggest that it may be useful for both public health research and environmental sustainability.
Limitations and Future Research Directions
This study has several limitations. The small sample size of human participants (n = 10) limits the statistical power and generalizability of the findings. Moreover, human gut microbiota is influenced by multiple factors that were not fully controlled in this short-term pilot study.
A key limitation is that diet was not systematically recorded or controlled. We did not use standardized tools such as food diaries or questionnaires, even though diet is known to strongly influence the gut microbiota21. As a result, some of the observed changes may be partly related to differences in participants’ eating habits rather than the probiotic intervention alone.
CFU-based methods capture only culturable bacteria and may not fully represent the diversity of microbial communities22,23. This limits the interpretation of overall microbiota composition. Furthermore, this method does not distinguish specific bacterial taxa and may include residual surface-associated bacteria despite sterilization, limiting interpretation of true gut colonization. Although 16S rRNA sequencing was used to analyze microbial composition, more detailed methods such as whole-genome or metagenomic sequencing were not included. These methods could provide deeper insight into microbial function and species-level differences24. Additionally, the absence of a placebo control group means that some reported effects, such as reduced bloating or fatigue, may be influenced by subjective bias. Another limitation is that detailed information about the probiotic formulations, including strain-level composition, CFU dosage, and manufacturer, was not available, which may limit the reproducibility of the intervention.
Detailed rearing conditions for the larvae, including diet, environmental factors, and housing conditions, were not strictly controlled or recorded, which may have influenced the observed microbial outcomes.
Although Hermetia illucens is a practical and ethical model, insect microbiota differ from those of humans in both composition and ecological structure13,14 , with some species exhibiting highly specialized and host-specific microbial communities25. This limits the extent to which findings can be directly translated to human health. Future studies should include additional model systems, such as mammals, along with larger-scale human trials to further investigate these findings.
In addition, the wide age range of participants (13–78 years) may have introduced biological variability, as gut microbiota composition and responses to probiotics can differ across age groups. This variability was not controlled for and should be considered when interpreting the results.
Finally, the saline control group also showed a substantial increase in CFU counts relative to baseline, suggesting that some of the observed changes may reflect time-dependent growth, developmental factors, or other experimental conditions rather than probiotic exposure alone.
Taken together, these limitations suggest that the findings should be viewed as exploratory rather than confirmatory.
Conclusion
This study found that probiotic supplementation was associated with changes in gut microbiota in human participants, while increases in culturable bacterial counts were observed in Hermetia illucens larvae under the experimental conditions. Human participants showed changes in microbiota scores and reported digestive benefits, while the insect model showed increased levels of culturable bacteria, including both probiotic-treated and control groups.
Although direct comparisons between species are limited, the insect model demonstrated measurable changes and may serve as a preliminary model for exploratory investigation of culture-detectable bacterial changes.
These findings suggest that insect systems may be used as a practical tool for early-stage investigation rather than direct comparison with human outcomes. Future studies with larger sample sizes and standardized analytical methods will be needed to further validate these findings.
Acknowledgements
The author would like to thank Yoon Young Kim, MD, PhD, from the College of Medicine at The Catholic University of Korea, for early advice during the study design process. The author also acknowledges Chanung Lee of Vanderbilt University for his valuable discussions and encouragement throughout this project.
References
- P. Hemarajata, J. Versalovic. Effects of probiotics on gut microbiota: mechanisms of intestinal immunomodulation and neuromodulation. Therapeutic Advances in Gastroenterology. Vol. 6, pg. 39–51, 2013, https://doi.org/10.1177/1756283X12459294. [↩] [↩]
- T. Zuo, S. C. Ng. The gut microbiota in the pathogenesis and therapeutics of inflammatory bowel disease. Frontiers in Microbiology. Vol. 9, pg. 2247, 2018, https://doi.org/10.3389/fmicb.2018.02247 [↩] [↩] [↩]
- J. Qin, Y. Li, Z. Cai, S. Li, J. Zhu, F. Zhang, S. Liang, W. Zhang, Y. Guan, D. Shen, Y. Peng, D. Zhang, Z. Jian, L. Chen, Y. Luo, Q. Li, Z. Wang, J. Zhang, G. Zhang, Y. Li, H. Wu, D. Qin, Y. Zhu, S. Liang, J. Yang, F. Wang, J. Li, X. Huang, J. Li, X. Li, L. Cao, B. Li, X. Li, Y. Fan, H. Li, X. Li, J. Liu, J. Liu, J. Li, Q. Xu, S. Wang, Y. Zhang, W. Zhang, Y. Zhao, J. Wang, H. Wang, H. Zheng, Y. Chen, J. Yang, M. Yang, J. Li, J. Wang, S. Zhou, H. Liu, Y. Zhang, D. Qin, S. Yang, M. Zhao, Z. Li, X. Li, X. Liu, Y. Zhang, X. Zhang, H. Zhang, Y. Zhang, H. Liu, J. Zhu, X. Li, Q. Zheng, Y. Wang, J. Hao, G. Li, X. Wang, S. Zhang, Y. Wang, H. Li, Z. Zhang, J. Wang, Y. Guo, X. Zhang, L. Zhao, J. Li, Y. Chen, J. Li, H. Zhang, J. Zhao, J. Liang, J. Zhang, Y. Wang, J. Wang, W. Li, J. Li. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature. Vol. 490, pg. 55–60, 2012, https://doi.org/10.1038/nature11450. [↩] [↩]
- P. J. Turnbaugh, R. E. Ley, M. A. Mahowald, V. Magrini, E. R. Mardis, J. I. Gordon. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. Vol. 444, pg. 1027–1031, 2006, https://doi.org/10.1038/nature05414. [↩] [↩]
- C. Hill, F. Guarner, G. Reid, G. R. Gibson, D. J. Merenstein, B. Pot, L. Morelli, R. Canani, H. J. Flint, S. Salminen, P. C. Calder, M. E. Sanders. Expert consensus document: the international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nature Reviews Gastroenterology & Hepatology. Vol. 11, pg. 506–514, 2014, https://doi.org/10.1038/nrgastro.2014.66. [↩] [↩] [↩] [↩]
- J. F. Cryan, T. G. Dinan. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nature Reviews Neuroscience. Vol. 13, pg. 701–712, 2012, https://doi.org/10.1038/nrn3346. [↩] [↩] [↩]
- E. D. Lewis, J. M. Antony, D. C. Crowley, A. A. Alkhabbaz, M. A. Evans. Efficacy of Lactobacillus paracasei HA-196 and Bifidobacterium longum R0175 in alleviating symptoms of irritable bowel syndrome (IBS): a randomized, placebo-controlled study. Nutrients. Vol. 12, pg. 1159, 2020, https://doi.org/10.3390/nu12041159. [↩]
- T. S. B. Schmidt, M. R. Hayward, L. P. Coelho, S. Li, M. Zeller, J. Sunagawa, P. Bork. Extensive transmission of microbes along the gastrointestinal tract. eLife. Vol. 8, pg. e42693, 2019, https://doi.org/10.7554/eLife.42693. [↩] [↩] [↩]
- S. Kitamoto, H. Nagao-Kitamoto, R. Hein, S. Shimada, T. Kitamoto, T. S. Miyoshi, H. Nakanishi, K. Morita, Y. Ishii, K. Sato. The bacterial connection between the oral cavity and gut diseases. Journal of Dental Research. Vol. 99, pg. 1021–1029, 2020, https://doi.org/10.1177/0022034520924633. [↩] [↩] [↩]
- Balls M. Replacement of animal procedures: alternatives in research. ALTEX. 2009;26(1):3–9. https://doi.org/10.14573/altex.2009.1.3 [↩]
- D. Bruno, M. Bonelli, F. De Filippis, C. Di Lelio, M. Tettamanti, M. Casartelli, G. Ercolini, S. Caccia. The intestinal microbiota of Hermetia illucens larvae is affected by diet and shows a diverse composition in the different midgut regions. Applied and Environmental Microbiology. Vol. 85, pg. e01864-18, 2019, https://doi.org/10.1128/AEM.01864-18. [↩] [↩]
- E. Wynants, L. Frooninckx, S. Crauwels, S. Verreth, K. De Smet, K. Sandrock, K. Wohlfahrt, L. Van Schelt, M. Depraetere, J. Lievens. Assessing the microbiota of black soldier fly larvae (Hermetia illucens) reared on organic waste streams. Microbial Ecology. Vol. 77, pg. 913–930, 2019, https://doi.org/10.1007/s00248-018-1286-x. [↩] [↩] [↩]
- K. Schmidt, P. Engel. Mechanisms underlying gut microbiota–host interactions in insects. Journal of Experimental Biology. Vol. 224, pg. jeb207696, 2021, https://doi.org/10.1242/jeb.207696. [↩] [↩]
- A. E. Douglas. Multiorganismal insects: diversity and function of resident microorganisms. Annual Review of Entomology. Vol. 60, pg. 17–34, 2015, https://doi.org/10.1146/annurev-ento-010814-020822. [↩] [↩]
- J. F. Cryan, K. J. O’Riordan, C. S. M. Cowan, K. V. Sandhu, T. F. S. Bastiaanssen, M. Boehme, M. G. Codagnone, S. Cussotto, C. Fulling, A. V. Golubeva, K. E. Guzzetta, M. Jaggar, C. M. Long-Smith, J. M. Lyte, J. A. Martin, A. Molinero-Perez, G. Moloney, E. Morelli, E. Morillas, R. O’Connor, J. S. Cruz-Pereira, V. L. Peterson, K. Rea, N. L. Ritz, E. Sherwin, S. Spichak, E. M. Teichman, M. van de Wouw, A. P. Ventura-Silva, S. E. Wallace-Fitzsimons, N. Hyland, G. Clarke, T. G. Dinan. The microbiota–gut–brain axis. Physiological Reviews. Vol. 99, pg. 1877–2013, 2019, https://doi.org/10.1152/physrev.00018.2018. [↩]
- G. Sharon, T. R. Sampson, D. H. Geschwind, S. K. Mazmanian. The central nervous system and the gut microbiome. Cell. Vol. 167, pg. 915–932, 2016, https://doi.org/10.1016/j.cell.2016.10.027. [↩]
- M. E. Sanders, D. J. Merenstein, G. Reid, G. R. Gibson, R. A. Rastall. Probiotics and prebiotics in intestinal health and disease: from biology to the clinic. Nature Reviews Gastroenterology & Hepatology. Vol. 16, pg. 605–616, 2019, https://doi.org/10.1038/s41575-019-0173-3. [↩]
- J. Suez, N. Zmora, E. Segal, E. Elinav. The pros, cons, and many unknowns of probiotics. Nature Medicine. Vol. 25, pg. 716–729, 2019, https://doi.org/10.1038/s41591-019-0439-x. [↩]
- A. D. Kostic, E. Chun, L. Robertson, C. A. Gallini, M. Michaud, T. E. Clancy, D. C. Chung, P. Lochhead, G. L. Hold, E. M. El-Omar, D. Brenner, C. S. Fuchs, M. Meyerson, W. S. Garrett. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host & Microbe. Vol. 14, pg. 207–215, 2013, https://doi.org/10.1016/j.chom.2013.07.007. [↩]
- S. Diener, C. Zurbrügg, K. Tockner. Conversion of organic material by black soldier fly larvae: establishing optimal feeding rates. Waste Management & Research. Vol. 27, pg. 603–610, 2009, https://doi.org/10.1177/0734242X09103838. [↩]
- N. Zmora, J. Suez, E. Elinav. You are what you eat: diet, health and the gut microbiota. Nature Reviews Gastroenterology & Hepatology. Vol. 16, pg. 35–56, 2019, https://doi.org/10.1038/s41575-018-0061-2. [↩]
- J. C. Lagier, P. Hugon, S. Khelaifia, P. E. Fournier, B. La Scola, D. Raoult. The rebirth of culture in microbiology through the example of culturomics to study human gut microbiota. Clinical Microbiology Reviews. Vol. 28, pg. 237–264, 2015, https://doi.org/10.1128/CMR.00014-14. [↩]
- J. C. Lagier, G. Dubourg, M. Million, F. Cadoret, M. Bilen, F. Fenollar, A. Levasseur, J. M. Rolain, P. E. Fournier, D. Raoult. Culturing the human microbiota and culturomics. Nature Reviews Microbiology. Vol. 16, pg. 540–550, 2018, https://doi.org/10.1038/s41579-018-0041-0. [↩]
- C. Quince, A. W. Walker, J. T. Simpson, N. J. Loman, N. Segata. Shotgun metagenomics, from sampling to analysis. Nature Biotechnology. Vol. 35, pg. 833–844, 2017, https://doi.org/10.1038/nbt.3935. [↩]
- P. Engel, N. A. Moran. Functional and evolutionary insights into the simple yet specific gut microbiota of the honey bee from metagenomic analysis. Gut Microbes. Vol. 4, pg. 60–65, 2013, https://doi.org/10.4161/gmic.22517. [↩]

 and Family-Integrated Care (FIC): Global Trends and Local Provider Awareness in Fresno County, California")

{kind=link}